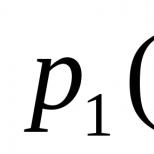Oppfanging og absorpsjon av store partikler av cellen. Ikke-cellulære strukturer
Vesikulær overføring kan deles inn i to typer: eksocytose - fjerning av makromolekylære produkter fra cellen, og endocytose - absorpsjon av makromolekyler av cellen.
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. Alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler kan gå inn i en slik primær vakuole, eller endosom, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet gjennom transmembranoverføring.
Den viktigste biologiske betydningen av endocytose er anskaffelsen av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med et lysosom, en vakuole som inneholder et sett med hydrolytiske enzymer.
Endocytose er formelt delt inn i pinocytose og fagocytose.
Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter
karakteristisk for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager). I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen
fagocytiske celler. Under fagocytose dannes store endocytiske vakuoler - fagosomer, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocytose
(pinocytose og fagocytose), såkalt fordi det går som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel,
partikler av sot eller fargestoffer.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, bretter, skiller små flytende medium volumer.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra cellemembranen.
overflaten og strekker seg dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Dette er de såkalte kantgropene. De kalles det fordi
På sidene av cytoplasmaet er plasmamembranen dekket, kledd, med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner grenser til og dekker små fremspring og groper. Disse hullene er
i nesten alle dyreceller opptar de omtrent 2 % av celleoverflaten. Grenselaget består hovedsakelig av klatrinproteinet assosiert med en rekke ekstra proteiner.
Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag langs omkretsen av det fremvoksende pinosomet.
Etter at den avgrensede vesikkelen skiller seg fra plasmolemmaet og begynner å bevege seg dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, og endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Reseptormediert endocytose. Effektiviteten av endocytose øker betydelig hvis den formidles av membranreseptorer som binder seg til molekyler av det absorberte stoffet eller molekyler som ligger på overflaten av det fagocyterte objektet - ligander (fra latin u^age - for å binde). Senere (etter absorpsjon av stoffet) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet. Et eksempel på en reseptormediert interaksjon er fagocytose av en bakteriell leukocytt.
Transcytose(fra lat. 1gash - gjennom, gjennom og gresk suYuz - celle) en prosess som er karakteristisk for noen typer celler, som kombinerer tegn på endocytose og eksocytose. En endocytisk vesikkel dannes på den ene celleoverflaten, som overføres til den motsatte celleoverflaten og som blir en eksocytisk vesikkel, frigjør innholdet til det ekstracellulære rommet.
Eksocytose
Plasmamembranen tar del i fjerning av stoffer fra cellen ved hjelp av eksocytose, en prosess som er det motsatte av endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster).
41 .Endoplasmatisk retikulum (retikulum).
I et lysmikroskop i fibriblaster etter fiksering og farging kan man se at periferien av cellene (ektoplasma) farges svakt, mens den sentrale delen av cellene (endoplasma) oppfatter fargestoffer godt. Så K. Porter i 1945 så i et elektronmikroskop at den endoplasmatiske sonen er fylt med et stort antall små vakuoler og kanaler som forbinder seg med hverandre og danner noe som et løst nettverk (retikulum). Det ble sett at stablene av disse vakuolene og tubuli var begrenset av tynne membraner. Så det ble oppdaget endoplasmatisk retikulum, eller endoplasmatisk retikulum. Senere, på 1950-tallet, ved hjelp av metoden med ultratynne seksjoner, var det mulig å belyse strukturen til denne formasjonen og å oppdage dens heterogenitet. Det viktigste viste seg å være at endoplasmatisk retikulum (ER) finnes i nesten alle eukaryoter.
En slik elektronmikroskopisk analyse gjorde det mulig å skille to typer ER: granulær (grov) og glatt.
1. Hooke oppdaget eksistensen av celler 2. Eksistensen av encellede organismer oppdaget Leeuwenhoek
4. Celler som inneholder en kjerne kalles eukaryoter
5. De strukturelle komponentene i en eukaryot celle inkluderer kjernen, ribosomer, plastider, mitokondrier, golgi-komplekset, det endoplasmatiske retikulum
6. Den intracellulære strukturen hvor den viktigste arvelige informasjonen er lagret kalles kjernen
7. Kjernen består av en kjernematrise og 2 membraner
8. Antall kjerner i en celle er vanligvis 1
9. Kompakt intranukleær struktur kalt kromatin
10. Den biologiske membranen som dekker hele cellen kalles cytoplasmatisk membran
11. Grunnlaget for alle biologiske membraner er polysakkarider
12. Biologiske membraner må inneholde proteiner
13. Et tynt lag med karbohydrater på den ytre overflaten av plasmamembranen kalles glykokalyxen
14. Hovedegenskapen til biologiske membraner er deres selektive permeabilitet
15. Planteceller er beskyttet av en membran, som består av cellulose
16. Absorpsjon av store partikler av en celle kalles fagocytose.
17. Absorpsjonen av væskedråper av cellen kalles pinocytose.
18. En del av en levende celle uten plasmamembran og kjerne kalles cytoplasma 19. Sammensetningen av cytoplasmaet inkluderer protoplasten og kjernen
20. Hovedstoffet i cytoplasmaet, oppløselig i vann, kalles glukose.
21. En del av cytoplasmaet, representert av støtte-kontraktile strukturer (komplekser), kalles vakuoler
22. Intracellulære strukturer som ikke er dens obligatoriske komponenter kalles inneslutninger
23. Ikke-membranorganeller som sørger for biosyntese av proteiner med en genetisk bestemt struktur kalles ribosomer.
24. Et komplett ribosom består av 2 underenheter
25. Sammensetningen av ribosomet inkluderer ... .
26. Hovedfunksjonen til ribosomer er proteinsyntese
27. Komplekser av ett molekyl av mRNA (mRNA) og dusinvis av ribosomer assosiert med det kalles ....
28. Grunnlaget for cellesenteret er mikrotubuli
29. En enkelt sentriol er ... .
30. Bevegelsesorganeller inkluderer flageller, flimmerhår
31. Systemet med tanker og tubuli koblet sammen til et enkelt intracellulært rom, avgrenset fra resten av cytoplasmaet av en lukket intracellulær membran, kalles EPS
32. Hovedfunksjonen til EPS er syntesen av organiske stoffer.
33. Ribosomer er lokalisert på overflaten av den grove ER
34. En del av endoplasmatisk retikulum, på overflaten som ribosomer befinner seg på, kalles grov EPS
35. Hovedfunksjonen til den granulære ER er syntesen av proteiner.
36. En del av det endoplasmatiske retikulumet, på overflaten som det ikke er ribosomer av, kalles glatte eps
37. Syntese av sukker og lipider skjer i hulrommet i den agranulære ER
38. Systemet med flate enkeltmembransisterner kalles Golgi-komplekset
39. Akkumulering av stoffer, deres modifisering og sortering, pakking av sluttprodukter i enkeltmembranvesikler, utskillelse av sekretoriske vakuoler utenfor cellen og dannelse av primære lysosomer - dette er funksjonene til Golgi-komplekset
40. Enkeltmembranvesikler som inneholder hydrolytiske enzymer kalles Goljilysosomkomplekset.
41. Væskefylte store enkeltmembranhulrom kalles vakuoler.
42. Innholdet av vakuoler kalles cellesaft
43. To-membranorganeller (som inkluderer ytre og indre membraner) inkluderer plastider og mitokondrier
44. Organeller som inneholder eget DNA, alle typer RNA, ribosomer og er i stand til å syntetisere noen proteiner er plastider og mitokondrier.
45. Hovedfunksjonen til mitokondrier er å skaffe energi i prosessen med cellulær respirasjon.
46. Hovedstoffet som er energikilden i cellen er ATP
Store molekyler av biopolymerer transporteres praktisk talt ikke gjennom membraner, og likevel kan de komme inn i cellen som følge av endocytose. Den er delt inn i fagocytose og pinocytose. Disse prosessene er assosiert med kraftig aktivitet og mobilitet av cytoplasma. Fagocytose er fangst og absorpsjon av store partikler av en celle (noen ganger til og med hele celler og deres deler). Fagocytose og pinocytose forløper veldig likt, derfor gjenspeiler disse konseptene bare forskjellen i volumene av absorberte stoffer. Felles for dem er at de absorberte stoffene på celleoverflaten er omgitt av en membran i form av en vakuole, som beveger seg inne i cellen (enten en fagocytisk eller pinocytisk vesikkel. Disse prosessene er forbundet med energiforbruk; opphør av ATP-syntese hemmer dem fullstendig. for eksempel veggene i tarmene, mange mikrovilli, betydelig øke overflaten som absorpsjon skjer gjennom. Plasmamembranen er også involvert i fjerning av stoffer fra cellen, dette skjer i prosessen eksocytose. Slik skilles hormoner, polysakkarider, proteiner, fettdråper og andre celleprodukter ut. De er innelukket i membranbundne vesikler og nærmer seg plasmalemmaet. Begge membranene smelter sammen og innholdet i vesikkelen slippes ut i miljøet rundt cellen.
Celler er også i stand til å absorbere makromolekyler og partikler ved hjelp av en lignende eksocytose mekanisme, men i omvendt rekkefølge. Absorbert materiale er gradvis omgitt av et lite område plasmamembran, som først invaginerer og deretter deler seg av og dannes intracellulær vesikkel som inneholder materiale fanget av cellen. Denne prosessen med dannelse av intracellulære vesikler rundt materialet som absorberes av cellen kalles endocytose.
Avhengig av størrelsen på vesiklene som dannes, skilles to typer endocytose:
1) pinocytose- absorpsjon av væske og oppløste stoffer gjennom små bobler, og
2) fagocytose- absorpsjon av store partikler som mikroorganismer eller celleavfall. I dette tilfellet dannes det store bobler, kalt vakuoler og absorpsjon av korpuskulært materiale: bakterier, store virus, døende egne celler i kroppen eller fremmede celler, som for eksempel erytrocytter av forskjellige typer, utføres av celler ( makrofager ,nøytrofiler)
Væske og oppløste stoffer tas kontinuerlig opp av de fleste celler gjennom pinocytose, mens store partikler hovedsakelig tas opp av spesialiserte celler - fagocytter. Derfor brukes begrepene "pinocytose" og "endocytose" vanligvis i samme betydning.
pinocytose karakterisert ved absorpsjon og intracellulær ødeleggelse av makromolekylære forbindelser, slik som proteiner og proteinkomplekser, nukleinsyrer, polysakkarider, lipoproteiner. Gjenstanden for pinocytose som en faktor for uspesifikk immunforsvar er spesielt giftstoffene til mikroorganismer. Adhesjon av stoffer på celleoverflaten fører til lokal invaginasjon (invaginasjon) av membranen, som kulminerer med dannelsen av en pinocytisk vesikkel av svært liten størrelse (ca. 0,1 mikron). Flere sammenslåtte bobler danner en større formasjon - pinosom. I neste trinn smelter pinosomene sammen med lysosomer inneholder hydrolytiske enzymer som bryter ned polymermolekyler til monomerer. I tilfeller der prosessen med pinocytose realiseres gjennom reseptorapparatet, i pinosomer, før fusjon med lysosomer, observeres løsrivelsen av fangede molekyler fra reseptorer, som, som en del av dattervesiklene, går tilbake til celleoverflaten.
Makromolekyler som proteiner, nukleinsyrer, polysakkarider, lipoproteinkomplekser og andre passerer ikke gjennom cellemembraner, i motsetning til hvordan ioner og monomerer transporteres. Transporten av mikromolekyler, deres komplekser, partikler inn og ut av cellen skjer på en helt annen måte - gjennom vesikulær overføring. Dette begrepet betyr at ulike makromolekyler, biopolymerer eller deres komplekser ikke kan komme inn i cellen gjennom plasmamembranen. Og ikke bare gjennom det: noen cellemembraner er ikke i stand til transmembranoverføring av biopolymerer, med unntak av membraner som har spesielle proteinkompleksbærere - poriner (membraner av mitokondrier, plastider, peroksisomer). Makromolekyler kommer inn i cellen eller fra ett membranrom til et annet innelukket i vakuoler eller vesikler. Slik vesikulær overføring kan deles inn i to typer: eksocytose- fjerning av makromolekylære produkter fra cellen, og endocytose- absorpsjon av makromolekyler av cellen (fig. 133).
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. I en slik primærvakuole, eller i endosom, kan alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler komme inn, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet ved transmembranoverføring. Den viktigste biologiske betydningen av endocytose er oppkjøpet av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med lysosomet, en vakuole som inneholder et sett med hydrolytiske enzymer (se nedenfor).
Endocytose er formelt delt inn i pinocytose og fagocytose(Fig. 134). Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter, er karakteristiske for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager). Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocyto h (pinocytose og fagocytose), såkalt fordi det fortsetter som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel sotpartikler eller fargestoffer.
Uspesifikk endocytose er ofte ledsaget av innledende sorpsjon av innfangningsmaterialet av plasmamembranens glykokalyx. Glykokalyxen, på grunn av de sure gruppene i polysakkaridene, har en negativ ladning og binder seg godt til forskjellige positivt ladede grupper av proteiner. Med slik adsorpsjon absorberes uspesifikk endocytose, makromolekyler og små partikler (sure proteiner, ferritin, antistoffer, virioner, kolloidale partikler). Væskefase pinocytose fører til absorpsjon sammen med det flytende mediet av løselige molekyler som ikke binder seg til plasmalemmaet.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, folder, og skiller små volumer av det flytende mediet (fig. 135, 136). Den første typen forekomst av en pinocytisk vesikkel, pinosomer, er typisk for celler i tarmepitel, endotel, for amøber, den andre - for fagocytter og fibroblaster. Disse prosessene er avhengige av tilførsel av energi: respirasjonshemmere blokkerer disse prosessene.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra celleoverflaten og går dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Disse er de såkalte avgrensede groper. De kalles det fordi plasmamembranen fra siden av cytoplasmaet er dekket, kledd med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner så å si grenser til, dekker små fremspring, groper (fig. 137). Nesten alle dyreceller har disse gropene; de opptar omtrent 2% av celleoverflaten. Omgivende lag består hovedsakelig av protein clathrin assosiert med en rekke ekstra proteiner. Tre molekyler av clathrin, sammen med tre molekyler av et lavmolekylært protein, danner strukturen til et triskelion, som ligner et tre-strålet hakekors (fig. 138). Clathrin triskelions på den indre overflaten av gropene i plasmamembranen danner et løst nettverk bestående av femkanter og sekskanter, vanligvis ligner en kurv. Klathrinlaget dekker hele omkretsen av de separerende primære endocytiske vakuolene, avgrenset av vesikler.
Clathrin tilhører en av de såkalte artene. "dressing" proteiner (COP - belagte proteiner). Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag rundt omkretsen av det fremvoksende pinosomet, den primære endosomale vesikkelen - den "kantede" vesikkelen. i separasjonen av det primære endosomet er også proteiner involvert - dynaminer, som polymeriserer rundt halsen på skillevesikkelen (fig. 139).
Etter at den kantede vesikkelen skiller seg fra plasmolemmaet og begynner å bli overført dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Det ble funnet at membranene i de avgrensede gropene inneholder relativt lite kolesterol, noe som kan bestemme reduksjonen i membranstivhet og bidra til dannelse av bobler. Den biologiske betydningen av utseendet til en clathrin "belegg" langs periferien av vesiklene kan være at den gir adhesjon av de avgrensede vesiklene til elementene i cytoskjelettet og deres påfølgende transport i cellen, og forhindrer dem i å smelte sammen med hverandre .
Intensiteten av væskefase uspesifikk pinocytose kan være svært høy. Så epitelcellen i tynntarmen danner opptil 1000 pinosomer per sekund, og makrofager danner omtrent 125 pinosomer per minutt. Størrelsen på pinosomer er liten, deres nedre grense er 60–130 nm, men deres overflod fører til det faktum at under endocytose blir plasmolemma raskt erstattet, som om "brukt" på dannelsen av mange små vakuoler. Så i makrofager erstattes hele plasmamembranen på 30 minutter, i fibroblaster - om to timer.
Den videre skjebnen til endosomer kan være annerledes, noen av dem kan gå tilbake til celleoverflaten og smelte sammen med den, men de fleste av dem går inn i prosessen med intracellulær fordøyelse. Primære endosomer inneholder for det meste fremmede molekyler fanget i det flytende mediet og inneholder ikke hydrolytiske enzymer. endosomer kan smelte sammen mens de øker i størrelse. De smelter deretter sammen med primære lysosomer (se nedenfor), som introduserer enzymer i endosomhulen som hydrolyserer forskjellige biopolymerer. Virkningen av disse lysosomale hydrolasene forårsaker intracellulær fordøyelse - nedbrytning av polymerer til monomerer.
Som allerede nevnt, under fagocytose og pinocytose, mister celler et stort område av plasmolemma (se makrofager), som imidlertid raskt gjenopprettes under membranresirkulering, på grunn av tilbakeføring av vakuoler og deres inkorporering i plasmolemma. Dette skyldes det faktum at små vesikler kan skilles fra endosomer eller vakuoler, samt fra lysosomer, som igjen smelter sammen med plasmamembranen. Med slik resirkulering oppstår en slags "shuttle"-overføring av membraner: plasmolemma - pinosome - vakuole - plasmolemma. Dette fører til restaurering av det opprinnelige området av plasmamembranen. Det ble funnet at med en slik retur, membranresirkulering, holdes alt absorbert materiale tilbake i det gjenværende endosomet.
Spesifikk eller reseptor-mediert endocytose har en rekke forskjeller fra uspesifikke. Hovedsaken er at molekyler absorberes som det er spesifikke reseptorer for på plasmamembranen som bare er assosiert med denne typen molekyler. Ofte kalles slike molekyler som binder seg til reseptorproteiner på overflaten av celler ligander.
Reseptormediert endocytose ble først beskrevet i akkumulering av proteiner i fugleoocytter. Proteiner av eggeplommegranulat, vitellogeniner, syntetiseres i ulike vev, men så kommer de inn i eggstokkene med blodstrømmen, hvor de binder seg til spesielle membranreseptorer av oocytter og kommer deretter inn i cellen ved hjelp av endocytose, hvor eggeplommegranuler avsettes.
Et annet eksempel på selektiv endocytose er transport av kolesterol inn i cellen. Dette lipidet syntetiseres i leveren og danner i kombinasjon med andre fosfolipider og et proteinmolekyl det såkalte. low-density lipoprotein (LDL), som skilles ut av leverceller og fraktes gjennom hele kroppen av sirkulasjonssystemet (fig. 140). Spesielle plasmamembranreseptorer diffust plassert på overflaten av forskjellige celler gjenkjenner proteinkomponenten i LDL og danner et spesifikt reseptor-ligandkompleks. Etter dette beveger et slikt kompleks seg til sonen med avgrensede groper og blir internalisert - omgitt av en membran og nedsenket dypt inn i cytoplasmaet. Det har blitt vist at mutante reseptorer kan binde LDL, men ikke akkumuleres i området med avgrensede groper. I tillegg til LDL-reseptorer er det funnet mer enn to dusin andre stoffer involvert i reseptorendocytose av forskjellige stoffer, som alle bruker samme internaliseringsvei gjennom de avgrensede gropene. Sannsynligvis er deres rolle i akkumulering av reseptorer: en og samme kantede grop kan samle rundt 1000 reseptorer av forskjellige klasser. I fibroblaster er imidlertid LDL-reseptorklynger lokalisert i sonen med avgrensede groper selv i fravær av en ligand i mediet.
Den videre skjebnen til den absorberte LDL-partikkelen er at den gjennomgår forfall i sammensetningen sekundært lysosom. Etter nedsenking i cytoplasmaet til en avgrenset vesikkel lastet med LDL, er det et raskt tap av clathrinlaget, membranvesikler begynner å smelte sammen med hverandre og danner et endosom - en vakuole som inneholder absorberte LDL-partikler som fortsatt er assosiert med reseptorer på membranoverflaten . Deretter oppstår dissosiasjonen av ligand-reseptorkomplekset, små vakuoler spaltes fra endosomet, hvis membraner inneholder frie reseptorer. Disse vesiklene resirkuleres, inkorporeres i plasmamembranen, og dermed går reseptorene tilbake til celleoverflaten. Skjebnen til LDL er at de etter fusjon med lysosomer hydrolyseres til fritt kolesterol, som kan inkorporeres i cellemembraner.
Endosomer kjennetegnes ved en lavere pH-verdi (pH 4-5), et surere miljø enn andre cellevakuoler. Dette skyldes tilstedeværelsen i deres membraner av protonpumpeproteiner som pumper inn hydrogenioner med samtidig forbruk av ATP (H + -avhengig ATPase). Det sure miljøet i endosomer spiller en kritisk rolle i dissosiasjonen av reseptorer og ligander. I tillegg er et surt miljø optimalt for aktivering av hydrolytiske enzymer i lysosomer, som aktiveres ved fusjon av lysosomer med endosomer og fører til dannelsen endolysosomer, hvor spaltningen av absorberte biopolymerer skjer.
I noen tilfeller er skjebnen til dissosierte ligander ikke relatert til lysosomal hydrolyse. Så i noen celler, etter binding av plasmolemma-reseptorer til visse proteiner, synker clathrin-belagte vakuoler inn i cytoplasmaet og overføres til et annet område av cellen, hvor de smelter sammen igjen med plasmamembranen, og de bundne proteinene dissosieres fra reseptorer. Slik foregår overføringen, transcytose, av enkelte proteiner gjennom veggen i endotelcellen fra blodplasmaet til det intercellulære miljøet (fig. 141). Et annet eksempel på transcytose er overføring av antistoffer. Så hos pattedyr kan antistoffene til moren overføres til ungen gjennom melk. I dette tilfellet forblir reseptor-antistoffkomplekset uendret i endosomet.
Fagocytose
Som allerede nevnt er fagocytose en variant av endocytose og er assosiert med cellens absorpsjon av store aggregater av makromolekyler opp til levende eller døde celler. I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen til fagocytiske celler. Under fagocytose dannes det store endocytiske vakuoler - fagosomet, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
På overflaten av celler som er i stand til fagocytose (hos pattedyr er disse nøytrofiler og makrofager), er det et sett med reseptorer som interagerer med ligandproteiner. Ved bakterielle infeksjoner binder antistoffer mot bakterielle proteiner seg til overflaten av bakterieceller, og danner et lag der Fc-områdene til antistoffene ser utover. Dette laget gjenkjennes av spesifikke reseptorer på overflaten av makrofager og nøytrofiler, og ved bindingsstedene begynner absorpsjonen av bakterien ved å omslutte den med plasmamembranen til cellen (fig. 142).
Eksocytose
Plasmamembranen er involvert i fjerning av stoffer fra cellen ved hjelp av eksocytose- den omvendte prosessen med endocytose (se fig. 133).
Ved eksocytose nærmer intracellulære produkter innelukket i vakuoler eller vesikler og separert fra hyaloplasmaet med en membran plasmamembranen. Ved deres kontaktpunkter smelter plasmamembranen og vakuolmembranen sammen, og boblen tømmes ut i miljøet. Ved hjelp av eksocytose oppstår prosessen med resirkulering av membraner involvert i endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster). På samme måte fjernes noen polysakkarider (hemicelluloser) som er involvert i dannelsen av cellevegger fra cytoplasmaet til planteceller.
De fleste utskilte stoffene brukes av andre celler i flercellede organismer (utskillelse av melk, fordøyelsessaft, hormoner, etc.). Men ofte skiller celler ut stoffer til egne behov. For eksempel utføres veksten av plasmamembranen ved å legge inn deler av membranen som en del av eksocytiske vakuoler, noen av elementene i glykokalyxen utskilles av cellen i form av glykoproteinmolekyler, etc.
Hydrolytiske enzymer isolert fra celler ved eksocytose kan sorberes i glykokalyxlaget og gi membranbundet ekstracellulær spaltning av ulike biopolymerer og organiske molekyler. Membran ikke-cellulær fordøyelse er av stor betydning for dyr. Det ble funnet at i tarmepitelet til pattedyr i området med den såkalte børstegrensen til det absorberende epitelet, som er spesielt rikt på glykokalyx, finnes det en enorm mengde forskjellige enzymer. Noen av disse enzymene er av bukspyttkjertelopprinnelse (amylase, lipaser, ulike proteinaser, etc.), og noen skilles ut av selve epitelcellene (eksohydrolaser, som bryter ned hovedsakelig oligomerer og dimerer med dannelse av transporterte produkter).
©2015-2019 nettsted
Alle rettigheter tilhører deres forfattere. Dette nettstedet krever ikke forfatterskap, men tilbyr gratis bruk.
Opprettelsesdato for side: 2016-04-15
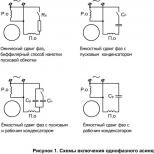
Vesikulær transport: endocytose og eksocytose
vesikulær overføring eksocytose endocytose
endosom
pinocytose og fagocytose
Uspesifikk endocyto
avgrensede groper clathrin
Spesifikk eller reseptor-mediert ligander.
sekundært lysosom
endolysosomer
Fagocytose
fagosomet fagolysosomer.
Eksocytose
eksocytose
Reseptorrollen til plasmalemmaet
Vi har allerede møtt denne funksjonen til plasmamembranen når vi ble kjent med dens transportfunksjoner. Bærerproteiner og pumper er også reseptorer som gjenkjenner og samhandler med visse ioner. Reseptorproteiner binder seg til ligander og deltar i utvalget av molekyler som kommer inn i celler.
Membranproteiner eller glykokalykselementer - glykoproteiner kan fungere som slike reseptorer på celleoverflaten. Slike følsomme steder for individuelle stoffer kan spres over overflaten av cellen eller samles i små soner.
Ulike celler av dyreorganismer kan ha forskjellige sett med reseptorer eller ulik følsomhet for samme reseptor.
Rollen til mange cellereseptorer er ikke bare i bindingen av spesifikke stoffer eller evnen til å reagere på fysiske faktorer, men også i overføringen av intercellulære signaler fra overflaten til cellen. For tiden er systemet for signaloverføring til celler ved hjelp av visse hormoner, som inkluderer peptidkjeder, blitt godt studert. Disse hormonene har vist seg å binde seg til spesifikke reseptorer på overflaten av cellens plasmamembran. Reseptorer, etter binding til hormonet, aktiverer et annet protein, som allerede er i den cytoplasmatiske delen av plasmamembranen, adenylatcyklase. Dette enzymet syntetiserer det sykliske AMP-molekylet fra ATP. Rollen til syklisk AMP (cAMP) er at det er en sekundær budbringer - en aktivator av enzymer - kinaser som forårsaker modifikasjoner av andre enzymproteiner. Så når bukspyttkjertelhormonet glukagon, produsert av A-celler på holmene i Langerhans, virker på levercellen, binder hormonet seg til en spesifikk reseptor, som stimulerer aktiveringen av adenylatcyklase. Syntetisert cAMP aktiverer proteinkinase A, som igjen aktiverer en kaskade av enzymer som til slutt bryter ned glykogen (dyrelagringspolysakkarid) til glukose. Virkningen av insulin er motsatt - det stimulerer inngangen av glukose til levercellene og dens avsetning i form av glykogen.
Generelt utfolder hendelseskjeden seg som følger: hormonet interagerer spesifikt med reseptordelen av dette systemet og, uten å trenge inn i cellen, aktiverer adenylatcyklase, som syntetiserer cAMP, som aktiverer eller hemmer et intracellulært enzym eller en gruppe enzymer . Dermed blir kommandoen, signalet fra plasmamembranen overført inne i cellen. Effektiviteten til dette adenylatcyklasesystemet er meget høy. Dermed kan samspillet mellom ett eller flere hormonmolekyler føre, på grunn av syntesen av mange cAMP-molekyler, til en signalforsterkning tusenvis av ganger. I dette tilfellet fungerer adenylatcyklasesystemet som en omformer av eksterne signaler.
Det er en annen måte som andre sekundære budbringere brukes på - dette er den såkalte. fosfatidylinositol-vei. Under påvirkning av et passende signal (noen nervemediatorer og proteiner) aktiveres enzymet fosfolipase C, som spalteripidet, som er en del av plasmamembranen. Hydrolyseproduktene av dette lipidet aktiverer på den ene siden proteinkinase C, som aktiverer kinasekaskaden, som fører til visse cellulære reaksjoner, og på den annen side fører til frigjøring av kalsiumioner, som regulerer en rekke cellulære prosesser.
Et annet eksempel på reseptoraktivitet er reseptorene for acetylkolin, en viktig nevrotransmitter. Acetylkolin, som frigjøres fra nerveenden, binder seg til reseptoren på muskelfiberen, forårsaker en impulsiv strøm av Na + inn i cellen (membrandepolarisering), og åpner umiddelbart rundt 2000 ionekanaler i området av den nevromuskulære enden.
Mangfoldet og spesifisiteten til settene av reseptorer på overflaten av celler fører til dannelsen av et veldig komplekst system av markører som gjør det mulig å skille ens egne celler (av samme individ eller av samme art) fra andres. Lignende celler inngår interaksjoner med hverandre, noe som fører til adhesjon av overflater (konjugering i protozoer og bakterier, dannelse av vevscellekomplekser). I dette tilfellet blir celler som er forskjellige i settet av determinantmarkører eller som ikke oppfatter dem, enten ekskludert fra slik interaksjon, eller blir ødelagt i høyere dyr som et resultat av immunologiske reaksjoner (se nedenfor).
Plasmamembranen er assosiert med lokalisering av spesifikke reseptorer som reagerer på fysiske faktorer. Så i plasmamembranen eller dens derivater i fotosyntetiske bakterier og blågrønne alger, er reseptorproteiner (klorofyller) som interagerer med lyskvanter lokalisert. I plasmamembranen til lysfølsomme dyreceller er det et spesielt system av fotoreseptorproteiner (rhodopsin), ved hjelp av hvilke lyssignalet omdannes til et kjemisk, som igjen fører til generering av en elektrisk impuls.
Intercellulær gjenkjennelse
I flercellede organismer, på grunn av intercellulære interaksjoner, dannes komplekse cellulære ensembler, hvis vedlikehold kan utføres på forskjellige måter. I germinalt, embryonalt vev, spesielt i de tidlige utviklingsstadiene, forblir celler koblet til hverandre på grunn av overflatenes evne til å holde sammen. Denne eiendommen vedheft(tilkobling, adhesjon) av celler kan bestemmes av egenskapene til overflaten deres, som spesifikt samhandler med hverandre. Mekanismen til disse forbindelsene er godt studert, den er gitt av interaksjonen mellom glykoproteiner i plasmamembraner. Med en slik intercellulær interaksjon av celler mellom plasmamembraner forblir det alltid et gap på omtrent 20 nm bredt, fylt med glykokalyx. Behandling av vev med enzymer som krenker integriteten til glykokalyxen (slim som virker hydrolytisk på muciner, mukopolysakkarider) eller skader plasmamembranen (proteaser) fører til isolering av celler fra hverandre, til deres dissosiasjon. Men hvis dissosiasjonsfaktoren fjernes, kan cellene settes sammen igjen og reaggregere. Så det er mulig å dissosiere celler av svamper i forskjellige farger, oransje og gul. Det viste seg at det dannes to typer aggregater i blandingen av disse cellene: de som kun består av gule og kun oransje celler. I dette tilfellet organiserer blandede cellesuspensjoner seg selv, og gjenoppretter den opprinnelige flercellede strukturen. Lignende resultater ble oppnådd med separerte cellesuspensjoner av amfibieembryoer; i dette tilfellet er det en selektiv romlig separasjon av ektodermceller fra endodermen og fra mesenkymet. Videre, hvis vev fra sene stadier av embryonal utvikling brukes til reaggregering, samles ulike celleensembler med vev- og organspesifisitet uavhengig i et reagensrør, epitelaggregater som ligner på nyretubuli, etc.
Det ble funnet at transmembrane glykoproteiner er ansvarlige for aggregeringen av homogene celler. Direkte for forbindelsen, vedheft, celler er ansvarlige for molekylene til den såkalte. CAM-proteiner (celleadhesjonsmolekyler). Noen av dem forbinder celler med hverandre på grunn av intermolekylære interaksjoner, andre danner spesielle intercellulære forbindelser eller kontakter.
Interaksjoner mellom adhesive proteiner kan være homofil når naboceller binder seg til hverandre ved hjelp av homogene molekyler, heterofile når forskjellige typer CAM-er på naboceller er involvert i adhesjon. Intercellulær binding skjer gjennom ytterligere linkermolekyler.
Det finnes flere klasser av CAM-proteiner. Dette er cadheriner, immunglobulinlignende N-CAM (nervecelleadhesjonsmolekyler), selektiner, integriner.
Cadheriner er integrerte fibrillære membranproteiner som danner parallelle homodimerer. Separate domener av disse proteinene er assosiert med Ca 2+ ioner, noe som gir dem en viss stivhet. Det er mer enn 40 arter av cadheriner. Dermed er E-cadherin karakteristisk for celler fra forhåndsimplanterte embryoer og epitelceller fra voksne organismer. P-cadherin er karakteristisk for trofoblast-, placenta- og epidermisceller; N-cadherin er lokalisert på overflaten av nerveceller, linseceller og på hjerte- og skjelettmuskulatur.
Adhesjonsmolekyler for nerveceller(N-CAM) tilhører immunoglobulin-superfamilien, de danner forbindelser mellom nerveceller. Noen av N-CAM-ene er involvert i koblingen av synapser, så vel som i adhesjonen av celler i immunsystemet.
selectins også, integrerte proteiner i plasmamembranen er involvert i adhesjonen av endotelceller, i bindingen av blodplater, leukocytter.
Integriner er heterodimerer, med a- og b-kjeder. Integriner forbinder først og fremst celler med ekstracellulære substrater, men de kan også delta i celleadhesjon til hverandre.
Gjenkjennelse av fremmede proteiner
Som allerede nevnt utvikler fremmede makromolekyler (antigener) som har kommet inn i kroppen en kompleks kompleks reaksjon - en immunreaksjon. Dens essens ligger i det faktum at noen av lymfocyttene produserer spesielle proteiner - antistoffer som spesifikt binder seg til antigener. For eksempel gjenkjenner makrofager antigen-antistoffkomplekser med deres overflatereseptorer og absorberer dem (for eksempel absorpsjon av bakterier under fagocytose).
I kroppen til alle virveldyr er det i tillegg et system for mottak av fremmede celler eller deres egne, men med endrede plasmamembranproteiner, for eksempel under virusinfeksjoner eller mutasjoner, ofte assosiert med tumordegenerasjon av celler.
Proteiner er lokalisert på overflaten av alle virveldyrceller, den såkalte. stort histokompatibilitetskompleks(hoved histokompatibilitetskompleks - MHC). Dette er integrerte proteiner, glykoproteiner, heterodimerer. Det er veldig viktig å huske at hvert individ har et annet sett av disse MHC-proteinene. Dette skyldes det faktum at de er veldig polymorfe, fordi hvert individ har et stort antall alternerende former av samme gen (mer enn 100), i tillegg er det 7-8 loci som koder for MHC-molekyler. Dette fører til det faktum at hver celle i en gitt organisme, som har et sett med MHC-proteiner, vil være forskjellig fra cellene til et individ av samme art. En spesiell form for lymfocytter, T-lymfocytter, gjenkjenner MHC i kroppen, men den minste endring i strukturen til MHC (for eksempel assosiasjon med et virus eller resultatet av en mutasjon i individuelle celler), fører til faktum at T-lymfocytter gjenkjenner slike endrede celler og ødelegger dem, men ikke ved fagocytose. De skiller ut spesifikke perforinproteiner fra sekretoriske vakuoler, som er innebygd i den cytoplasmatiske membranen til den endrede cellen, danner transmembrankanaler i den, noe som gjør plasmamembranen permeabel, noe som fører til at den endrede cellen dør (fig. 143, 144).
Spesielle intercellulære forbindelser
I tillegg til disse relativt enkle adhesive (men spesifikke) bindingene (fig. 145), er det en rekke spesielle intercellulære strukturer, kontakter eller forbindelser som utfører visse funksjoner. Dette er låse-, forankrings- og kommunikasjonsforbindelser (fig. 146).
Låse eller tett forbindelse karakteristisk for enkeltlags epitel. Dette er sonen hvor de ytre lagene av de to plasmamembranene er så nærme som mulig. Trelagsmembranen sees ofte i denne kontakten: de to ytre osmofile lagene til begge membranene ser ut til å smelte sammen til ett felles lag 2–3 nm tykt. Fusjon av membraner skjer ikke over hele området med tett kontakt, men er en serie av punktkonvergens av membraner (fig. 147a, 148).
På plane preparater av plasmamembranbrudd i sonen med tett kontakt, ved bruk av fryse- og flismetoden, ble det funnet at kontaktpunktene til membranene er rader med kuler. Dette er proteinene occludin og claudin, spesielle integrerte proteiner i plasmamembranen, bygget i rader. Slike rader med kuler eller strimler kan krysse hverandre på en slik måte at de så å si danner et gitter eller nettverk på spalteflaten. Denne strukturen er veldig typisk for epitel, spesielt kjertel og tarm. I det siste tilfellet danner tett kontakt en kontinuerlig sone for fusjon av plasmamembraner, som omgir cellen i dens apikale (øvre, ser inn i tarmlumen) del (fig. 148). Dermed er hver celle i laget så å si omgitt av et bånd av denne kontakten. Slike strukturer kan også sees med spesielle flekker i et lysmikroskop. De fikk navnet fra morfologer endeplater. Det viste seg at i dette tilfellet er rollen til den lukkende tette kontakten ikke bare i den mekaniske forbindelsen av celler med hverandre. Dette kontaktområdet er dårlig permeabelt for makromolekyler og ioner, og dermed låser det, blokkerer de intercellulære hulrommene, og isolerer dem (og med dem det indre miljøet i kroppen) fra det ytre miljøet (i dette tilfellet tarmlumen).
Dette kan demonstreres ved bruk av elektrontette kontrastmidler som lantanhydroksidløsning. Hvis lumen i tarmen eller kanalen til en kjertel er fylt med en løsning av lantanhydroksid, så på seksjoner under et elektronmikroskop, har sonene der dette stoffet befinner seg en høy elektrontetthet og vil være mørke. Det viste seg at verken sonen med tett kontakt eller de intercellulære rommene under den mørkere. Hvis tight junctions er skadet (ved lett enzymatisk behandling eller fjerning av Ca ++ ioner), trenger lantan også inn i de intercellulære områdene. Tilsvarende har tight junctions vist seg å være ugjennomtrengelige for hemoglobin og ferritin i tubuli i nyrene.
1. Hooke oppdaget eksistensen av celler 2. Eksistensen av encellede organismer oppdaget Leeuwenhoek
4. Celler som inneholder en kjerne kalles eukaryoter
5. De strukturelle komponentene i en eukaryot celle inkluderer kjernen, ribosomer, plastider, mitokondrier, golgi-komplekset, det endoplasmatiske retikulum
6. Den intracellulære strukturen hvor den viktigste arvelige informasjonen er lagret kalles kjernen
7. Kjernen består av en kjernematrise og 2 membraner
8. Antall kjerner i en celle er vanligvis 1
9. Kompakt intranukleær struktur kalt kromatin
10. Den biologiske membranen som dekker hele cellen kalles cytoplasmatisk membran
11. Grunnlaget for alle biologiske membraner er polysakkarider
12. Biologiske membraner må inneholde proteiner
13. Et tynt lag med karbohydrater på den ytre overflaten av plasmamembranen kalles glykokalyxen
14. Hovedegenskapen til biologiske membraner er deres selektive permeabilitet
15. Planteceller er beskyttet av en membran, som består av cellulose
16. Absorpsjon av store partikler av en celle kalles fagocytose.
17. Absorpsjonen av væskedråper av cellen kalles pinocytose.
18. En del av en levende celle uten plasmamembran og kjerne kalles cytoplasma 19. Sammensetningen av cytoplasmaet inkluderer protoplasten og kjernen
20. Hovedstoffet i cytoplasmaet, oppløselig i vann, kalles glukose.
21. En del av cytoplasmaet, representert av støtte-kontraktile strukturer (komplekser), kalles vakuoler
22. Intracellulære strukturer som ikke er dens obligatoriske komponenter kalles inneslutninger
23. Ikke-membranorganeller som sørger for biosyntese av proteiner med en genetisk bestemt struktur kalles ribosomer.
24. Et komplett ribosom består av 2 underenheter
25. Sammensetningen av ribosomet inkluderer ... .
26. Hovedfunksjonen til ribosomer er proteinsyntese
27. Komplekser av ett molekyl av mRNA (mRNA) og dusinvis av ribosomer assosiert med det kalles ....
28. Grunnlaget for cellesenteret er mikrotubuli
29. En enkelt sentriol er ... .
30. Bevegelsesorganeller inkluderer flageller, flimmerhår
31. Systemet med tanker og tubuli koblet sammen til et enkelt intracellulært rom, avgrenset fra resten av cytoplasmaet av en lukket intracellulær membran, kalles EPS
32. Hovedfunksjonen til EPS er syntesen av organiske stoffer.
33. Ribosomer er lokalisert på overflaten av den grove ER
34. En del av endoplasmatisk retikulum, på overflaten som ribosomer befinner seg på, kalles grov EPS
35. Hovedfunksjonen til den granulære ER er syntesen av proteiner.
36. En del av det endoplasmatiske retikulumet, på overflaten som det ikke er ribosomer av, kalles glatte eps
37. Syntese av sukker og lipider skjer i hulrommet i den agranulære ER
38. Systemet med flate enkeltmembransisterner kalles Golgi-komplekset
39. Akkumulering av stoffer, deres modifisering og sortering, pakking av sluttprodukter i enkeltmembranvesikler, utskillelse av sekretoriske vakuoler utenfor cellen og dannelse av primære lysosomer - dette er funksjonene til Golgi-komplekset
40. Enkeltmembranvesikler som inneholder hydrolytiske enzymer kalles Goljilysosomkomplekset.
41. Væskefylte store enkeltmembranhulrom kalles vakuoler.
42. Innholdet av vakuoler kalles cellesaft
43. To-membranorganeller (som inkluderer ytre og indre membraner) inkluderer plastider og mitokondrier
44. Organeller som inneholder eget DNA, alle typer RNA, ribosomer og er i stand til å syntetisere noen proteiner er plastider og mitokondrier.
45. Hovedfunksjonen til mitokondrier er å skaffe energi i prosessen med cellulær respirasjon.
46. Hovedstoffet som er energikilden i cellen er ATP
Store molekyler av biopolymerer transporteres praktisk talt ikke gjennom membraner, og likevel kan de komme inn i cellen som følge av endocytose. Det er delt inn i fagocytose og pinocytose. Disse prosessene er assosiert med kraftig aktivitet og mobilitet av cytoplasma. Fagocytose er fangst og absorpsjon av store partikler av en celle (noen ganger til og med hele celler og deres deler). Fagocytose og pinocytose forløper veldig likt, derfor gjenspeiler disse konseptene bare forskjellen i volumene av absorberte stoffer. Felles for dem er at de absorberte stoffene på celleoverflaten er omgitt av en membran i form av en vakuole, som beveger seg inne i cellen (eller fagocytisk eller pinocytisk vesikkel, fig. 19). Disse prosessene er forbundet med energiforbruk; opphør av ATP-syntese hemmer dem fullstendig. På overflaten av epitelceller som fôrer, for eksempel veggene i tarmen, er mange mikrovilli synlige, noe som betydelig øker overflaten som absorpsjon skjer gjennom. Plasmamembranen er også involvert i fjerning av stoffer fra cellen, dette skjer i prosessen med eksocytose. Slik skilles hormoner, polysakkarider, proteiner, fettdråper og andre celleprodukter ut. De er innelukket i membranbundne vesikler og nærmer seg plasmalemmaet. Begge membranene smelter sammen og innholdet i vesikkelen slippes ut i miljøet rundt cellen.
Celler er også i stand til å absorbere makromolekyler og partikler ved å bruke en mekanisme som ligner på eksocytose, men i omvendt rekkefølge. Det absorberte stoffet omgis gradvis av et lite område av plasmamembranen, som først invaginerer og deretter splittes av, og danner en intracellulær vesikkel som inneholder materialet som fanges opp av cellen (fig. 8-76). Denne prosessen med dannelse av intracellulære vesikler rundt materialet som absorberes av cellen kalles endocytose.
Avhengig av størrelsen på vesiklene som dannes, skilles to typer endocytose:
Væske og oppløste stoffer tas kontinuerlig opp av de fleste celler gjennom pinocytose, mens store partikler hovedsakelig tas opp av spesialiserte celler, fagocytter. Derfor brukes begrepene "pinocytose" og "endocytose" vanligvis i samme betydning.
Pinocytose er preget av absorpsjon og intracellulær ødeleggelse av makromolekylære forbindelser som proteiner og proteinkomplekser, nukleinsyrer, polysakkarider, lipoproteiner. Gjenstanden for pinocytose som en faktor for uspesifikk immunforsvar er spesielt giftstoffene til mikroorganismer.
På fig. B.1 viser de påfølgende stadiene av fangst og intracellulær fordøyelse av løselige makromolekyler lokalisert i det ekstracellulære rommet (endocytose av makromolekyler av fagocytter). Adhesjonen av slike molekyler på cellen kan utføres på to måter: ikke-spesifikk - som et resultat av et tilfeldig møte av molekyler med cellen, og spesifikk, som avhenger av eksisterende reseptorer på overflaten av den pinocytiske cellen . I sistnevnte tilfelle fungerer ekstracellulære stoffer som ligander som interagerer med de tilsvarende reseptorene.
Adhesjon av stoffer på celleoverflaten fører til lokal invaginasjon (invaginasjon) av membranen, som kulminerer med dannelsen av en pinocytisk vesikkel av svært liten størrelse (ca. 0,1 mikron). Flere sammenvoksede vesikler danner en større formasjon - pinosomet. På neste trinn smelter pinosomer sammen med lysosomer som inneholder hydrolytiske enzymer som bryter ned polymermolekyler til monomerer. I tilfeller der prosessen med pinocytose realiseres gjennom reseptorapparatet, i pinosomer, før fusjon med lysosomer, observeres løsrivelsen av fangede molekyler fra reseptorer, som, som en del av dattervesiklene, går tilbake til celleoverflaten.
Del 3. Transmembranbevegelse av makromolekyler
Makromolekyler kan transporteres over plasmamembranen. Prosessen der celler tar inn store molekyler kalles endocytose. Noen av disse molekylene (for eksempel polysakkarider, proteiner og polynukleotider) tjener som en kilde til næringsstoffer. Endocytose gjør det også mulig å regulere innholdet av visse membrankomponenter, spesielt hormonreseptorer. Endocytose kan brukes til å studere cellulære funksjoner mer detaljert. Celler av en type kan transformeres med en annen type DNA og dermed endre funksjon eller fenotype.
I slike eksperimenter brukes ofte spesifikke gener, noe som gir en unik mulighet til å studere mekanismene for deres regulering. Transformasjonen av celler ved hjelp av DNA utføres ved endocytose - dette er måten DNA kommer inn i cellen. Transformasjon utføres vanligvis i nærvær av kalsiumfosfat, siden Ca 2+ stimulerer endocytose og DNA-utfelling, noe som letter dens inntreden i cellen ved endocytose.
Makromolekyler forlater cellen eksocytose. Ved både endocytose og eksocytose dannes det vesikler som smelter sammen med plasmamembranen eller løsner fra den.
3.1. Endocytose: typer endocytose og mekanisme
Alle eukaryote celler en del av plasmamembranen er hele tiden inne i cytoplasmaet. Dette skjer som et resultat invaginasjon av et fragment av plasmamembranen, utdanning endocytisk vesikkel , lukke halsen på vesikkelen og snøre den inn i cytoplasmaet sammen med innholdet (Fig. 18). Deretter kan vesikler smelte sammen med andre membranstrukturer og dermed overføre innholdet til andre cellulære rom eller til og med tilbake til det ekstracellulære rommet. De fleste endocytiske vesikler smelter sammen med primære lysosomer og danner sekundære lysosomer, som inneholder hydrolytiske enzymer og er spesialiserte organeller. Makromolekyler fordøyes i dem til aminosyrer, enkle sukkerarter og nukleotider, som diffunderer fra vesiklene og utnyttes i cytoplasmaet.
For endocytose trenger du:
1) energi, kilden som vanligvis er ATP;
2) ekstracellulært Ca 2+;
3) kontraktile elementer i cellen(sannsynligvis mikrofilamentsystemer).
Endocytose kan deles inn tre hovedtyper:
1. Fagocytose kun utført involverer spesialiserte celler (Fig. 19), slik som makrofager og granulocytter. Under fagocytose absorberes store partikler - virus, bakterier, celler eller deres fragmenter. Makrofager er eksepsjonelt aktive i denne forbindelse og kan skru på et volum på 25 % av sitt eget volum på 1 time. Dette internaliserer 3 % av plasmamembranen hvert minutt, eller hele membranen hvert 30. minutt.
 2. pinocytose finnes i alle celler. Med den, cellen absorberer væsker
og komponenter oppløst i den (fig. 20). Flytende fase pinocytose er ikke-selektiv prosess
, hvor mengden av det oppløste stoffet absorbert i sammensetningen av vesiklene ganske enkelt er proporsjonal med konsentrasjonen i den ekstracellulære væsken. Slike vesikler dannes utelukkende aktivt. For eksempel, i fibroblaster, er internaliseringshastigheten av plasmamembranen 1/3 av hastigheten som er karakteristisk for makrofager. I dette tilfellet forbrukes membranen raskere enn den syntetiseres. Samtidig endres ikke cellens overflate og volum mye, noe som indikerer at membranen gjenopprettes på grunn av eksocytose eller på grunn av dens re-inklusjon i samme hastighet som den konsumeres.
2. pinocytose finnes i alle celler. Med den, cellen absorberer væsker
og komponenter oppløst i den (fig. 20). Flytende fase pinocytose er ikke-selektiv prosess
, hvor mengden av det oppløste stoffet absorbert i sammensetningen av vesiklene ganske enkelt er proporsjonal med konsentrasjonen i den ekstracellulære væsken. Slike vesikler dannes utelukkende aktivt. For eksempel, i fibroblaster, er internaliseringshastigheten av plasmamembranen 1/3 av hastigheten som er karakteristisk for makrofager. I dette tilfellet forbrukes membranen raskere enn den syntetiseres. Samtidig endres ikke cellens overflate og volum mye, noe som indikerer at membranen gjenopprettes på grunn av eksocytose eller på grunn av dens re-inklusjon i samme hastighet som den konsumeres.
 3. Reseptormediert endocytose(reopptak av nevrotransmitter) - endocytose, der membranreseptorer binder seg til molekyler av det absorberte stoffet, eller molekyler lokalisert på overflaten av det fagocyterte objektet - ligander (fra lat. ligare–binde(Fig. 21) )
. Senere (etter absorpsjon av et stoff eller objekt) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet.
3. Reseptormediert endocytose(reopptak av nevrotransmitter) - endocytose, der membranreseptorer binder seg til molekyler av det absorberte stoffet, eller molekyler lokalisert på overflaten av det fagocyterte objektet - ligander (fra lat. ligare–binde(Fig. 21) )
. Senere (etter absorpsjon av et stoff eller objekt) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet.
Et eksempel på reseptormediert endocytose er fagocytose av en bakterie med en leukocytt. Siden leukocyttens plasmolemma har reseptorer for immunglobuliner (antistoffer), øker fagocytosehastigheten hvis overflaten av bakteriecelleveggen er dekket med antistoffer (opsoniner - fra det greske opson–krydder).
 Reseptormediert endocytose er en aktiv spesifikk prosess der cellemembranen buler inn i cellen og danner avgrensede groper
. Den intracellulære siden av den avgrensede gropen inneholder sett med adaptive proteiner
(adaptin, clathrin, som bestemmer den nødvendige krumningen av bulen, og andre proteiner) (Fig. 22). Når liganden er bundet fra miljøet som omgir cellen, danner de avgrensede gropene intracellulære vesikler (kantede vesikler). Reseptormediert endocytose slås på for raskt og kontrollert opptak av cellen av den aktuelle liganden. Disse vesiklene mister raskt grensen og smelter sammen med hverandre, og danner større vesikler - endosomer.
Reseptormediert endocytose er en aktiv spesifikk prosess der cellemembranen buler inn i cellen og danner avgrensede groper
. Den intracellulære siden av den avgrensede gropen inneholder sett med adaptive proteiner
(adaptin, clathrin, som bestemmer den nødvendige krumningen av bulen, og andre proteiner) (Fig. 22). Når liganden er bundet fra miljøet som omgir cellen, danner de avgrensede gropene intracellulære vesikler (kantede vesikler). Reseptormediert endocytose slås på for raskt og kontrollert opptak av cellen av den aktuelle liganden. Disse vesiklene mister raskt grensen og smelter sammen med hverandre, og danner større vesikler - endosomer.
clathrin- intracellulært protein, hovedkomponenten i membranen til avgrensede vesikler dannet under reseptorendocytose (fig. 23).
 Tre clathrin-molekyler er assosiert med hverandre i den C-terminale enden på en slik måte at clathrin-trimeren har form som en triskelion. Som et resultat av polymerisering danner clathrin et lukket tredimensjonalt nettverk som ligner en fotball. Størrelsen på clathrin vesikler er omtrent 100 nm.
Tre clathrin-molekyler er assosiert med hverandre i den C-terminale enden på en slik måte at clathrin-trimeren har form som en triskelion. Som et resultat av polymerisering danner clathrin et lukket tredimensjonalt nettverk som ligner en fotball. Størrelsen på clathrin vesikler er omtrent 100 nm.
Avgrensede groper kan okkupere opptil 2% av overflaten til noen celler. Endocytiske vesikler som inneholder lipoproteiner med lav tetthet (LDL) og deres reseptorer smelter sammen med lysosomer i cellen. Reseptorene frigjøres og returneres til overflaten av cellemembranen, og LDL-apoproteinet spaltes og den tilsvarende kolesterolesteren metaboliseres. Syntese av LDL-reseptorer reguleres av sekundære eller tertiære produkter av pinocytose, dvs. stoffer som dannes under metabolismen av LDL, slik som kolesterol.
3.2. Eksocytose: kalsiumavhengig og kalsiumuavhengig.
De fleste celler frigjør makromolekyler til miljøet ved eksocytose . Denne prosessen spiller også en rolle i membranfornyelse når dets komponenter syntetisert i Golgi-apparatet leveres som en del av vesikler til plasmamembranen (fig. 24).
 |
Ris. 24. Sammenligning av mekanismene for endocytose og eksocytose.
Mellom ekso- og endocytose, i tillegg til forskjellen i bevegelsesretningen til stoffer, er det en annen betydelig forskjell: når eksocytose fortsette fusjon av to indre cytoplasmatiske monolag , mens kl endocyose de ytre monolagene smelter sammen.
Stoffer frigjort ved eksocytose, kan deles i tre kategorier:
1) stoffer som binder seg til celleoverflaten og å bli perifere proteiner, slik som antigener;
2) stoffer som inngår i den ekstracellulære matrisen f.eks. kollagen og glykosaminoglykaner;
3) stoffer som slippes ut i det ekstracellulære miljøet og fungerer som signalmolekyler for andre celler.
Eukaryoter er utmerkede to typer eksocytose:
1. Kalsiumuavhengig konstitutiv eksocytose forekommer i nesten alle eukaryote celler. Det er en nødvendig prosess å bygge en ekstracellulær matrise og levere proteiner til den ytre cellemembranen. I dette tilfellet blir sekretoriske vesikler levert til celleoverflaten og smelter sammen med den ytre membranen når de dannes.
2. kalsiumavhengig ikke-konstitutiv eksocytose forekommer, for eksempel, i kjemiske synapser eller celler som produserer makromolekylære hormoner. Denne eksocytosen tjener f.eks. å isolere nevrotransmittere. Ved denne typen eksocytose akkumuleres sekretoriske vesikler i cellen, og prosessen med utgivelsen utløses av et visst signal mediert av en rask økning i konsentrasjonen kalsiumioner i cytosolen til cellen. I presynaptiske membraner utføres prosessen av et spesielt kalsiumavhengig proteinkompleks SNARE.
Makromolekyler som proteiner, nukleinsyrer, polysakkarider, lipoproteinkomplekser og andre passerer ikke gjennom cellemembraner, i motsetning til hvordan ioner og monomerer transporteres. Transporten av mikromolekyler, deres komplekser, partikler inn og ut av cellen skjer på en helt annen måte - gjennom vesikulær overføring. Dette begrepet betyr at ulike makromolekyler, biopolymerer eller deres komplekser ikke kan komme inn i cellen gjennom plasmamembranen. Og ikke bare gjennom det: noen cellemembraner er ikke i stand til transmembranoverføring av biopolymerer, med unntak av membraner som har spesielle proteinkompleksbærere - poriner (membraner av mitokondrier, plastider, peroksisomer). Makromolekyler kommer inn i cellen eller fra ett membranrom til et annet innelukket i vakuoler eller vesikler. Slik vesikulær overføring kan deles inn i to typer: eksocytose- fjerning av makromolekylære produkter fra cellen, og endocytose- absorpsjon av makromolekyler av cellen (fig. 133).
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. I en slik primærvakuole, eller i endosom, kan alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler komme inn, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet ved transmembranoverføring. Den viktigste biologiske betydningen av endocytose er oppkjøpet av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med lysosomet, en vakuole som inneholder et sett med hydrolytiske enzymer (se nedenfor).
Endocytose er formelt delt inn i pinocytose og fagocytose(Fig. 134). Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter, er karakteristiske for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager). Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocyto h (pinocytose og fagocytose), såkalt fordi det fortsetter som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel sotpartikler eller fargestoffer.
Uspesifikk endocytose er ofte ledsaget av innledende sorpsjon av innfangningsmaterialet av plasmamembranens glykokalyx. Glykokalyxen, på grunn av de sure gruppene i polysakkaridene, har en negativ ladning og binder seg godt til forskjellige positivt ladede grupper av proteiner. Med slik adsorpsjon absorberes uspesifikk endocytose, makromolekyler og små partikler (sure proteiner, ferritin, antistoffer, virioner, kolloidale partikler). Væskefase pinocytose fører til absorpsjon sammen med det flytende mediet av løselige molekyler som ikke binder seg til plasmalemmaet.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, folder, og skiller små volumer av det flytende mediet (fig. 135, 136). Den første typen forekomst av en pinocytisk vesikkel, pinosomer, er typisk for celler i tarmepitel, endotel, for amøber, den andre - for fagocytter og fibroblaster. Disse prosessene er avhengige av tilførsel av energi: respirasjonshemmere blokkerer disse prosessene.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra celleoverflaten og går dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Disse er de såkalte avgrensede groper. De kalles det fordi plasmamembranen fra siden av cytoplasmaet er dekket, kledd med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner så å si grenser til, dekker små fremspring, groper (fig. 137). Nesten alle dyreceller har disse gropene; de opptar omtrent 2% av celleoverflaten. Omgivende lag består hovedsakelig av protein clathrin assosiert med en rekke ekstra proteiner. Tre molekyler av clathrin, sammen med tre molekyler av et lavmolekylært protein, danner strukturen til et triskelion, som ligner et tre-strålet hakekors (fig. 138). Clathrin triskelions på den indre overflaten av gropene i plasmamembranen danner et løst nettverk bestående av femkanter og sekskanter, vanligvis ligner en kurv. Klathrinlaget dekker hele omkretsen av de separerende primære endocytiske vakuolene, avgrenset av vesikler.
Clathrin tilhører en av de såkalte artene. "dressing" proteiner (COP - belagte proteiner). Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag rundt omkretsen av det fremvoksende pinosomet, den primære endosomale vesikkelen - den "kantede" vesikkelen. i separasjonen av det primære endosomet er også proteiner involvert - dynaminer, som polymeriserer rundt halsen på skillevesikkelen (fig. 139).
Etter at den kantede vesikkelen skiller seg fra plasmolemmaet og begynner å bli overført dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Det ble funnet at membranene i de avgrensede gropene inneholder relativt lite kolesterol, noe som kan bestemme reduksjonen i membranstivhet og bidra til dannelse av bobler. Den biologiske betydningen av utseendet til en clathrin "belegg" langs periferien av vesiklene kan være at den gir adhesjon av de avgrensede vesiklene til elementene i cytoskjelettet og deres påfølgende transport i cellen, og forhindrer dem i å smelte sammen med hverandre .
Intensiteten av væskefase uspesifikk pinocytose kan være svært høy. Så epitelcellen i tynntarmen danner opptil 1000 pinosomer per sekund, og makrofager danner omtrent 125 pinosomer per minutt. Størrelsen på pinosomer er liten, deres nedre grense er 60–130 nm, men deres overflod fører til det faktum at under endocytose blir plasmolemma raskt erstattet, som om "brukt" på dannelsen av mange små vakuoler. Så i makrofager erstattes hele plasmamembranen på 30 minutter, i fibroblaster - om to timer.
Den videre skjebnen til endosomer kan være annerledes, noen av dem kan gå tilbake til celleoverflaten og smelte sammen med den, men de fleste av dem går inn i prosessen med intracellulær fordøyelse. Primære endosomer inneholder for det meste fremmede molekyler fanget i det flytende mediet og inneholder ikke hydrolytiske enzymer. endosomer kan smelte sammen mens de øker i størrelse. De smelter deretter sammen med primære lysosomer (se nedenfor), som introduserer enzymer i endosomhulen som hydrolyserer forskjellige biopolymerer. Virkningen av disse lysosomale hydrolasene forårsaker intracellulær fordøyelse - nedbrytning av polymerer til monomerer.
Som allerede nevnt, under fagocytose og pinocytose, mister celler et stort område av plasmolemma (se makrofager), som imidlertid raskt gjenopprettes under membranresirkulering, på grunn av tilbakeføring av vakuoler og deres inkorporering i plasmolemma. Dette skyldes det faktum at små vesikler kan skilles fra endosomer eller vakuoler, samt fra lysosomer, som igjen smelter sammen med plasmamembranen. Med slik resirkulering oppstår en slags "shuttle"-overføring av membraner: plasmolemma - pinosome - vakuole - plasmolemma. Dette fører til restaurering av det opprinnelige området av plasmamembranen. Det ble funnet at med en slik retur, membranresirkulering, holdes alt absorbert materiale tilbake i det gjenværende endosomet.
Spesifikk eller reseptor-mediert endocytose har en rekke forskjeller fra uspesifikke. Hovedsaken er at molekyler absorberes som det er spesifikke reseptorer for på plasmamembranen som bare er assosiert med denne typen molekyler. Ofte kalles slike molekyler som binder seg til reseptorproteiner på overflaten av celler ligander.
Reseptormediert endocytose ble først beskrevet i akkumulering av proteiner i fugleoocytter. Proteiner av eggeplommegranulat, vitellogeniner, syntetiseres i ulike vev, men så kommer de inn i eggstokkene med blodstrømmen, hvor de binder seg til spesielle membranreseptorer av oocytter og kommer deretter inn i cellen ved hjelp av endocytose, hvor eggeplommegranuler avsettes.
Et annet eksempel på selektiv endocytose er transport av kolesterol inn i cellen. Dette lipidet syntetiseres i leveren og danner i kombinasjon med andre fosfolipider og et proteinmolekyl det såkalte. low-density lipoprotein (LDL), som skilles ut av leverceller og fraktes gjennom hele kroppen av sirkulasjonssystemet (fig. 140). Spesielle plasmamembranreseptorer diffust plassert på overflaten av forskjellige celler gjenkjenner proteinkomponenten i LDL og danner et spesifikt reseptor-ligandkompleks. Etter dette beveger et slikt kompleks seg til sonen med avgrensede groper og blir internalisert - omgitt av en membran og nedsenket dypt inn i cytoplasmaet. Det har blitt vist at mutante reseptorer kan binde LDL, men ikke akkumuleres i området med avgrensede groper. I tillegg til LDL-reseptorer er det funnet mer enn to dusin andre stoffer involvert i reseptorendocytose av forskjellige stoffer, som alle bruker samme internaliseringsvei gjennom de avgrensede gropene. Sannsynligvis er deres rolle i akkumulering av reseptorer: en og samme kantede grop kan samle rundt 1000 reseptorer av forskjellige klasser. I fibroblaster er imidlertid LDL-reseptorklynger lokalisert i sonen med avgrensede groper selv i fravær av en ligand i mediet.
Den videre skjebnen til den absorberte LDL-partikkelen er at den gjennomgår forfall i sammensetningen sekundært lysosom. Etter nedsenking i cytoplasmaet til en avgrenset vesikkel lastet med LDL, er det et raskt tap av clathrinlaget, membranvesikler begynner å smelte sammen med hverandre og danner et endosom - en vakuole som inneholder absorberte LDL-partikler som fortsatt er assosiert med reseptorer på membranoverflaten . Deretter oppstår dissosiasjonen av ligand-reseptorkomplekset, små vakuoler spaltes fra endosomet, hvis membraner inneholder frie reseptorer. Disse vesiklene resirkuleres, inkorporeres i plasmamembranen, og dermed går reseptorene tilbake til celleoverflaten. Skjebnen til LDL er at de etter fusjon med lysosomer hydrolyseres til fritt kolesterol, som kan inkorporeres i cellemembraner.
Endosomer kjennetegnes ved en lavere pH-verdi (pH 4-5), et surere miljø enn andre cellevakuoler. Dette skyldes tilstedeværelsen i deres membraner av protonpumpeproteiner som pumper inn hydrogenioner med samtidig forbruk av ATP (H + -avhengig ATPase). Det sure miljøet i endosomer spiller en kritisk rolle i dissosiasjonen av reseptorer og ligander. I tillegg er et surt miljø optimalt for aktivering av hydrolytiske enzymer i lysosomer, som aktiveres ved fusjon av lysosomer med endosomer og fører til dannelsen endolysosomer, hvor spaltningen av absorberte biopolymerer skjer.
I noen tilfeller er skjebnen til dissosierte ligander ikke relatert til lysosomal hydrolyse. Så i noen celler, etter binding av plasmolemma-reseptorer til visse proteiner, synker clathrin-belagte vakuoler inn i cytoplasmaet og overføres til et annet område av cellen, hvor de smelter sammen igjen med plasmamembranen, og de bundne proteinene dissosieres fra reseptorer. Slik foregår overføringen, transcytose, av enkelte proteiner gjennom veggen i endotelcellen fra blodplasmaet til det intercellulære miljøet (fig. 141). Et annet eksempel på transcytose er overføring av antistoffer. Så hos pattedyr kan antistoffene til moren overføres til ungen gjennom melk. I dette tilfellet forblir reseptor-antistoffkomplekset uendret i endosomet.
Fagocytose
Som allerede nevnt er fagocytose en variant av endocytose og er assosiert med cellens absorpsjon av store aggregater av makromolekyler opp til levende eller døde celler. I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen til fagocytiske celler. Under fagocytose dannes det store endocytiske vakuoler - fagosomet, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
På overflaten av celler som er i stand til fagocytose (hos pattedyr er disse nøytrofiler og makrofager), er det et sett med reseptorer som interagerer med ligandproteiner. Ved bakterielle infeksjoner binder antistoffer mot bakterielle proteiner seg til overflaten av bakterieceller, og danner et lag der Fc-områdene til antistoffene ser utover. Dette laget gjenkjennes av spesifikke reseptorer på overflaten av makrofager og nøytrofiler, og ved bindingsstedene begynner absorpsjonen av bakterien ved å omslutte den med plasmamembranen til cellen (fig. 142).
Eksocytose
Plasmamembranen er involvert i fjerning av stoffer fra cellen ved hjelp av eksocytose- den omvendte prosessen med endocytose (se fig. 133).
Ved eksocytose nærmer intracellulære produkter innelukket i vakuoler eller vesikler og separert fra hyaloplasmaet med en membran plasmamembranen. Ved deres kontaktpunkter smelter plasmamembranen og vakuolmembranen sammen, og boblen tømmes ut i miljøet. Ved hjelp av eksocytose oppstår prosessen med resirkulering av membraner involvert i endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster). På samme måte fjernes noen polysakkarider (hemicelluloser) som er involvert i dannelsen av cellevegger fra cytoplasmaet til planteceller.
De fleste utskilte stoffene brukes av andre celler i flercellede organismer (utskillelse av melk, fordøyelsessaft, hormoner, etc.). Men ofte skiller celler ut stoffer til egne behov. For eksempel utføres veksten av plasmamembranen ved å legge inn deler av membranen som en del av eksocytiske vakuoler, noen av elementene i glykokalyxen utskilles av cellen i form av glykoproteinmolekyler, etc.
Hydrolytiske enzymer isolert fra celler ved eksocytose kan sorberes i glykokalyxlaget og gi membranbundet ekstracellulær spaltning av ulike biopolymerer og organiske molekyler. Membran ikke-cellulær fordøyelse er av stor betydning for dyr. Det ble funnet at i tarmepitelet til pattedyr i området med den såkalte børstegrensen til det absorberende epitelet, som er spesielt rikt på glykokalyx, finnes det en enorm mengde forskjellige enzymer. Noen av disse enzymene er av bukspyttkjertelopprinnelse (amylase, lipaser, ulike proteinaser, etc.), og noen skilles ut av selve epitelcellene (eksohydrolaser, som bryter ned hovedsakelig oligomerer og dimerer med dannelse av transporterte produkter).
©2015-2019 nettsted
Alle rettigheter tilhører deres forfattere. Dette nettstedet krever ikke forfatterskap, men tilbyr gratis bruk.
Opprettelsesdato for side: 2016-04-15
dobbel membran
Enkel membran
Lipoprotein
Selektiv transport av stoffer inn i en celle mot en konsentrasjonsgradient med energiforbruk
Inntrengning i cellen av stoffer langs konsentrasjonsgradienten uten energiforbruk
Bevegelse av fettuløselige stoffer gjennom ionekanaler i membranen
aktiv transport
K-na pumpe
cytoplasmatisk membran
Intracellulære fibrillære strukturer
Intercellulær gjenkjennelse
pinocytose
Fagocytose
Eksocytose
3.20. Oppfanging og absorpsjon av store partikler av en celle kalles:
2. Eksocytose
3. Endocytose
4. Pinocytose
3.21. Oppfanging og absorpsjon av en væske og stoffer oppløst i den av en celle kalles:
1. Fagocytose
2. Eksocytose
3. Endocytose
3.22. Karbohydratkjeder i glykokalyxen til dyreceller gir:
1. Fangst og absorpsjon
2. Beskyttelse mot utenlandske agenter
3. Sekresjon
3.23. Den mekaniske stabiliteten til plasmamembranen bestemmes av
1. Karbohydrater
3.24. Konstansen til celleformen sikres ved:
2. Cellevegg
3. Vakuoler
4. Flytende cytoplasma
3,25. Energi er nødvendig når stoffer kommer inn i cellen ved hjelp av:
1. Diffusjon
2. Tilrettelagt diffusjon
3,26. Energiforbruk oppstår ikke når stoffer kommer inn i cellen ved
1. Fago- og pinocytose
2. Endocytose og eksocytose
3. Passiv transport
4. Aktiv transport
3,27. Na, K, Ca-ioner kommer inn i cellen gjennom
1. Diffusjon
2. Tilrettelagt diffusjon
3,28. Tilrettelagt diffusjon er
1. Innfanging av flytende stoffer av cellemembranen og deres inntreden i cellens cytoplasma
2. Innfanging av faste partikler av cellemembranen og deres inntreden i cytoplasma
4. Bevegelse av stoffer over en membran mot en konsentrasjonsgradient
3.29. Passiv transport er
3. Selektiv transport av stoffer inn i en celle mot en konsentrasjonsgradient med energiforbruk
3.30 Aktiv transport er
1. Innfanging av flytende stoffer av cellemembranen og deres overføring til cellens cytoplasma
2. Innfanging av faste partikler av cellemembranen og deres overføring til cytoplasma
4. Inntreden i cellen av stoffer langs konsentrasjonsgradienten uten energiforbruk
3,31. Cellemembraner er et kompleks:
2. Nukleoprotein
3. Glykolipid
4. Glykoprotein
3,32. Celleorganellen - Golgi-apparatet er:
1. Ikke-membran
3. Dobbel membran
4. Spesiell
3,33. Celleorganellen, mitokondriet, er:
1. Ikke-membran
2. Enkel membran
4. Spesiell
3,34. Celleorganell - cellesenter er:
2. Enkel membran
3. Dobbel membran
4. Spesiell
3,35. Syntese skjer på grov EPS:
1. Lipider
Oppfanging og absorpsjon av store partikler av cellen kalles. Membrantransport av makromolekyler og partikler: endocytose og eksocytose (fagocytose og pinocytose)
Store molekyler av biopolymerer transporteres praktisk talt ikke gjennom membraner, og likevel kan de komme inn i cellen som følge av endocytose. Det er delt inn i fagocytose og pinocytose. Disse prosessene er assosiert med kraftig aktivitet og mobilitet av cytoplasma. Fagocytose er fangst og absorpsjon av store partikler av en celle (noen ganger til og med hele celler og deres deler). Fagocytose og pinocytose forløper veldig likt, derfor gjenspeiler disse konseptene bare forskjellen i volumene av absorberte stoffer. Felles for dem er at de absorberte stoffene på celleoverflaten er omgitt av en membran i form av en vakuole, som beveger seg inne i cellen (eller fagocytisk eller pinocytisk vesikkel, fig. 19). Disse prosessene er forbundet med energiforbruk; opphør av ATP-syntese hemmer dem fullstendig. På overflaten av epitelceller som fôrer, for eksempel veggene i tarmen, er mange mikrovilli synlige, noe som betydelig øker overflaten som absorpsjon skjer gjennom. Plasmamembranen er også involvert i fjerning av stoffer fra cellen, dette skjer i prosessen med eksocytose. Slik skilles hormoner, polysakkarider, proteiner, fettdråper og andre celleprodukter ut. De er innelukket i membranbundne vesikler og nærmer seg plasmalemmaet. Begge membranene smelter sammen og innholdet i vesikkelen slippes ut i miljøet rundt cellen.
Celler er også i stand til å absorbere makromolekyler og partikler ved å bruke en mekanisme som ligner på eksocytose, men i omvendt rekkefølge. Det absorberte stoffet omgis gradvis av et lite område av plasmamembranen, som først invaginerer og deretter splittes av, og danner en intracellulær vesikkel som inneholder materialet som fanges opp av cellen (fig. 8-76). Denne prosessen med dannelse av intracellulære vesikler rundt materialet som absorberes av cellen kalles endocytose.
Avhengig av størrelsen på vesiklene som dannes, skilles to typer endocytose:
Væske og oppløste stoffer tas kontinuerlig opp av de fleste celler gjennom pinocytose, mens store partikler hovedsakelig tas opp av spesialiserte celler, fagocytter. Derfor brukes begrepene "pinocytose" og "endocytose" vanligvis i samme betydning.
Pinocytose er preget av absorpsjon og intracellulær ødeleggelse av makromolekylære forbindelser som proteiner og proteinkomplekser, nukleinsyrer, polysakkarider, lipoproteiner. Gjenstanden for pinocytose som en faktor for uspesifikk immunforsvar er spesielt giftstoffene til mikroorganismer.
På fig. B.1 viser de påfølgende stadiene av fangst og intracellulær fordøyelse av løselige makromolekyler lokalisert i det ekstracellulære rommet (endocytose av makromolekyler av fagocytter). Adhesjonen av slike molekyler på cellen kan utføres på to måter: ikke-spesifikk - som et resultat av et tilfeldig møte av molekyler med cellen, og spesifikk, som avhenger av eksisterende reseptorer på overflaten av den pinocytiske cellen . I sistnevnte tilfelle fungerer ekstracellulære stoffer som ligander som interagerer med de tilsvarende reseptorene.
Adhesjon av stoffer på celleoverflaten fører til lokal invaginasjon (invaginasjon) av membranen, som kulminerer med dannelsen av en pinocytisk vesikkel av svært liten størrelse (ca. 0,1 mikron). Flere sammenvoksede vesikler danner en større formasjon - pinosomet. På neste trinn smelter pinosomer sammen med lysosomer som inneholder hydrolytiske enzymer som bryter ned polymermolekyler til monomerer. I tilfeller der prosessen med pinocytose realiseres gjennom reseptorapparatet, i pinosomer, før fusjon med lysosomer, observeres løsrivelsen av fangede molekyler fra reseptorer, som, som en del av dattervesiklene, går tilbake til celleoverflaten.
Ikke-cellulære strukturer
I dyrekroppen, i tillegg til individuelle celler, er det også ikke-cellulære strukturer som er sekundære til celler.
Ikke-cellulære strukturer er delt inn i:
1) kjernefysisk; 2) ikke-atomkraft
Kjernefysisk- inneholder en kjerne og oppstår ved cellefusjon, eller som et resultat av ufullstendig deling. Disse formasjonene inkluderer: symplaster og syncytia.
FRA implaster– Dette er store formasjoner som består av cytoplasma og et stort antall kjerner. Et eksempel på symplaster er skjelettmuskulatur, det ytre laget av placenta trofoblasten.
syncytium eller menigheter disse formasjonene er preget av det faktum at etter delingen av den opprinnelige cellen, forblir de nydannede cellene sammenkoblet av cytoplasmatiske broer. En slik midlertidig struktur oppstår under utviklingen av mannlige kjønnsceller, når delingen av cellekroppen ikke er fullstendig fullført.
Ikke-atomkraft– Dette er ikke-cellulære strukturer som representerer produktet av den vitale aktiviteten til individuelle grupper av celler. Et eksempel på slike strukturer er fibre og hovedstoffet (amorfe) i bindevevet, som produseres av fibroblastceller. Analoger av hovedstoffet er blodplasma og den flytende delen av lymfen.
Det må understrekes at det også finnes kjernefysiske celler i kroppen. Disse elementene inkorporerer cellemembranen og cytoplasmaet, er utstyrt med begrensede funksjoner og har mistet evnen til å reprodusere seg selv på grunn av fraværet av en kjerne. den erytrocytter og blodplater.
Generell plan for cellestrukturen
En eukaryot celle har 3 hovedkomponenter:
1. Cellemembran; 2. Cytoplasmer; 3. Kjerner.
Celleveggen avgrenser cellens cytoplasma fra miljøet eller fra naboceller.
Cytoplasma på sin side består den av hyaloplasma og organiserte strukturer, som inkluderer organeller og inneslutninger.
Cellekjernen har en kjernemembran, karyoplasma, kromatin (kromosomer), nukleolus.
Alle de listede komponentene til celler, som samhandler med hverandre, utfører funksjonene for å sikre eksistensen av cellen som helhet.
SKJEMA 1. Strukturelle komponenter i cellen
CELLEKONVOLUTTE
Celleveggen(plasmolemma) - er en perifer overflatestruktur som begrenser cellen fra utsiden og gir dens direkte forbindelse med det ekstracellulære miljøet, og derfor med alle stoffer og faktorer som påvirker cellen.
Struktur
Cellemembranen består av 3 lag (fig. 1):
1) ytre (supra-membran) lag - glycocalyx (Glicocalyx);
2) den faktiske membranen (biologisk membran);
3) submembranplate (kortikalt lag av plasmalemma).
Glycocalyx- dannes av glykoprotein- og glykolipidkomplekser assosiert med plasmalemmaet, som inkluderer forskjellige karbohydrater. Karbohydrater er lange, forgrenede kjeder av polysakkarider som er assosiert med proteiner og lipider som er en del av plasmalemmaet. Tykkelsen på glykokalyxen er 3-4 nm; den er iboende i nesten alle celler av animalsk opprinnelse, men med varierende alvorlighetsgrad. Polysakkaridkjedene til glykokalyxen er et slags apparat der celler gjensidig gjenkjenner og samhandler med mikromiljøet.
Egen membran(biologisk membran). Den strukturelle organiseringen av en biologisk membran er mest reflektert i Singer-Nikolsky-væske-mosaikkmodellen, ifølge hvilken fosfolipidmolekyler kommer i kontakt med deres hydrofobe ender (haler), og frastøter med hydrofile ender (hoder), danner et kontinuerlig dobbeltlag.
Helt integrerte proteiner er nedsenket i bilipidlaget (disse er hovedsakelig glykoproteiner), semi-integrerte proteiner er delvis nedsenket. Disse to gruppene av proteiner i det bilipide laget av membranen er plassert på en slik måte at deres ikke-polare deler er inkludert i dette laget av membranen på lokaliseringsstedene til de hydrofobe områdene av lipider (haler). Den polare delen av proteinmolekylet samhandler med lipidhodene som vender mot den vandige fasen.
I tillegg ligger en del av proteinene på overflaten av bilipidlaget, disse er de såkalte membranbundne eller perifere eller adsorberte proteinene.
Plasseringen av proteinmolekyler er ikke strengt begrenset, og avhengig av cellens funksjonelle tilstand kan deres gjensidige bevegelse i planet til bilipidlaget oppstå.
Slik variasjon i posisjonen til proteiner, og topografien til mikromolekylære komplekser av celleoverflaten, lik en mosaikk, ga navnet til væske-mosaikkmodellen til en biologisk membran.
Labiliteten (mobiliteten) til plasmamembranstrukturene avhenger av innholdet av kolesterolmolekyler i sammensetningen. Jo mer kolesterol som er inneholdt i membranen, desto lettere skjer bevegelsen av makromolekylære proteiner i bilipidlaget. Tykkelsen på den biologiske membranen er 5-7 nm.
undermembranplate(kortikalt lag) dannes av den tetteste delen av cytoplasmaet, rik på mikrofilamenter og mikrotubuli, som danner et høyt organisert nettverk, med deltakelse som de integrerte proteinene i plasmolemma beveger seg, cellens cytoskjelett- og bevegelsesfunksjoner er gitt , og eksocytoseprosesser realiseres. Tykkelsen på dette laget er omtrent 1 nm.
Funksjoner
Hovedfunksjonene utført av cellemembranen inkluderer følgende:
1) avgrensning;
2) transport av stoffer;
3) mottak;
4) sikre intercellulære kontakter.
Avgrensning og transport av metabolitter
Takket være differensieringen med miljøet beholder cellen sin individualitet, takket være transport kan cellen leve og fungere. Begge disse funksjonene er gjensidig utelukkende og komplementære til hverandre, og begge prosessene er rettet mot å opprettholde konstansen til egenskapene til det indre miljøet - cellehomeostase.
Transport fra miljøet inn i cellen kan være aktiv og passiv.
· Gjennom aktiv transport overføres mange organiske forbindelser mot en tetthetsgradient med energiforbruk på grunn av ATP-splitting, med deltagelse av enzymatiske transportsystemer.
· Passiv transport utføres ved diffusjon og gir overføring av vann, ioner, noen lavmolekylære forbindelser.
Transport av stoffer fra miljøet inn i cellen kalles endocytose, kalles prosessen med å fjerne stoffer fra cellen eksocytose.
Endocytose delt på fagocytose og pinocytose.
Fagocytose- dette er cellens fangst og absorpsjon av store partikler (bakterier, fragmenter av andre celler).
pinocytose- dette er fangst av mikromolekylære forbindelser som er i oppløst tilstand (væsker).
Endocytose fortsetter i flere påfølgende stadier:
1) Sorpsjon- overflaten av membranen av absorberte stoffer, hvis binding til plasmamembranen bestemmes av tilstedeværelsen av reseptormolekyler på overflaten.
2) Invaginasjon av plasmalemmaet inn i cellen. Til å begynne med ser invaginasjonene ut som åpne avrundede vesikler eller dype intussusceptions.
3) Løsning av invaginasjoner fra plasmalemmaet. De separerte vesiklene er fritt plassert i cytoplasmaet under plasmalemmaet. Bobler kan smelte sammen med hverandre.
4) Splitting av absorberte partikler ved hjelp av hydrolytiske enzymer som kommer fra lysosomer.
Noen ganger er det også en slik variant når en partikkel absorberes av en celleoverflate og passerer gjennom cytoplasmaet inn i miljøet til biomembranen og skilles ut fra cellen uforandret på den motsatte celleoverflaten. Et slikt fenomen kalles cytopempisom.
Eksocytose– Dette er fjerning av celleavfallsprodukter fra cytoplasma.
Det finnes flere typer eksocytose:
1) sekresjon;
2) utskillelse;
3) rekreasjon;
4) klasmatose.
Sekresjon- frigjøring av cellen av produktene av dens syntetiske aktivitet, nødvendig for å sikre de fysiologiske funksjonene til organene og systemene i kroppen.
Utskillelse- frigjøring av giftige metabolske produkter som utskilles utenfor kroppen.
rekreasjon- fjerning fra cellen av forbindelser som ikke endrer deres kjemiske struktur i prosessen med intracellulær metabolisme (vann, mineralsalter).
klasmatose- fjerning utenfor cellen av dens individuelle strukturelle komponenter.
Eksocytose består av en rekke påfølgende stadier:
1) akkumulering av produkter av den syntetiske aktiviteten til cellen i form av akkumuleringer omgitt av en biomembran som en del av sekkene og vesiklene til Golgi-komplekset;
2) bevegelsen av disse ansamlingene fra de sentrale områdene av cytoplasma til periferien;
3) inkorporering av sekkens biomembran i plasmalemmaet;
4) evakuering av innholdet i sekken inn i det intercellulære rommet.
resepsjon
Persepsjon (mottak) av cellen av ulike stimuli i mikromiljøet utføres med deltakelse av spesielle reseptorproteiner av plasmalemma. Spesifisiteten (selektiviteten) av interaksjonen mellom reseptorproteinet med en viss stimulus bestemmes av karbohydratkomponenten som er en del av dette proteinet. Overføring av det mottatte signalet til reseptoren inne i cellen kan utføres gjennom adenylatcyklasesystemet, som er en av dens veier.
Det skal bemerkes at komplekse mottaksprosesser er grunnlaget for gjensidig anerkjennelse av celler og derfor er en grunnleggende nødvendig betingelse for eksistensen av flercellede organismer.
Intercellulære kontakter (tilkoblinger)
Forbindelsen mellom celler i vev og organer til flercellede dyreorganismer er dannet av komplekse spesielle strukturer kalt intercellulære kontakter.
Strukturerte intercellulære kontakter er spesielt uttalt i det integumentære kantvevet, i epitelet.
Alle intercellulære kontakter er delt inn i tre grupper i henhold til deres funksjonelle formål:
1) intercellulære adhesjonskontakter (klebende);
2) isolerende;
3) kommunikasjon.
~Den første gruppen inkluderer: a) en enkel kontakt, b) en lås-type kontakt, c) en desmosom.
· Enkel kontakt- dette er konvergensen av plasmalemmaet til naboceller i en avstand på 15-20 nm. Fra siden av cytoplasmaet grenser ingen spesielle strukturer til denne sonen av membranen. En variant av enkel kontakt er interdigitering.
· Kontakt etter låstype- dette er et fremspring av overflaten av plasmamembranen til en celle inn i invaginatet (fremspringet) til en annen. Rollen til den tette lukkeforbindelsen er å mekanisk koble cellene til hverandre. Denne typen intercellulære forbindelser er karakteristisk for mange epitel, der den forbinder celler til et enkelt lag, noe som letter deres mekaniske festing til hverandre.
Det intermembrane (intercellulære) rommet og cytoplasmaet i "låse"-sonen har de samme egenskapene som i sonene med enkel kontakt med en avstand på 10-20 nm.
· Desmosomt er et lite område på opptil 0,5 µm i diameter, hvor et område med høy elektrontetthet er plassert mellom membranene, noen ganger med et lagdelt utseende. En del av elektrontett stoff grenser til plasmamembranen i området av desmosomet fra siden av cytoplasmaet slik at det indre laget av membranen ser ut til å være fortykket. Under fortykkelsen er et område med tynne fibriller som kan legges inn i en relativt tett matrise. Disse fibrillene danner ofte løkker og går tilbake til cytoplasmaet. Tynnere filamenter, som stammer fra tette plater i det nærmembranøse cytoplasmaet, passerer inn i det intercellulære rommet, hvor de danner et sentralt tett lag. Disse "intermembrane ligamentene" gir en direkte mekanisk forbindelse mellom nettverk av tonofilamenter av tilstøtende epitelceller eller andre celler.
~ Den andre gruppen inkluderer:
a) tett kontakt.
· Tett(lukke)kontakt er en sone hvor de ytre lagene til to plasmamembraner er så nærme som mulig. Trelagsmembranen sees ofte i denne kontakten: de to ytre osmiofile lagene til begge membranene ser ut til å smelte sammen til ett felles lag 2–3 nm tykt. Fusjon av membraner skjer ikke over hele området med tett kontakt, men er en serie punktkonvergens av membraner. Det er fastslått at kontaktpunktene til membranene er kuler av spesielle integrerte proteiner arrangert i rader. Disse rekkene med kuler kan krysse hverandre, slik at de så å si danner et gitter eller nettverk. Fra siden av cytoplasmaet i denne sonen er det mange fibriller med en diameter på 7 nm, som er plassert parallelt med plasmolemma. Kontaktområdet er ugjennomtrengelig for makromolekyler og ioner, og dermed låser, blokkerer de intercellulære hulrommene, og isolerer dem fra det ytre miljøet. Denne strukturen er typisk for epitel, spesielt for mage eller tarm.
~ Den tredje gruppen inkluderer:
a) gapkontakt (nexus).
· Gap kontakter- dette er kommunikasjonsforbindelser mellom celler gjennom spesielle proteinkomplekser - sammenhenger, som er involvert i direkte overføring av kjemikalier fra celle til celle.
Sonen til en slik forbindelse har dimensjoner på 0,5-3 μm og avstanden mellom plasmamembranene i dette området er 2-3 nm. I sonen til denne kontakten er partikler arrangert sekskantet - konnexoner med en diameter på 7-8 nm og en kanal i midten med en bredde på 1,5 nm. Connexon er sammensatt av seks underenheter av connectinproteinet. Connexons er bygget inn i membranen på en slik måte at de penetrerer den gjennom og gjennom, sammenfallende på plasmamembranene til to naboceller, lukker de ende mot ende. Som et resultat etableres en direkte kjemisk binding mellom cytoplasmaene til cellene. Denne typen kontakt er typisk for alle typer vev.
Vesikulær overføring kan deles inn i to typer: eksocytose - fjerning av makromolekylære produkter fra cellen, og endocytose - absorpsjon av makromolekyler av cellen.
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. Alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler kan gå inn i en slik primær vakuole, eller endosom, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet gjennom transmembranoverføring.
Den viktigste biologiske betydningen av endocytose er anskaffelsen av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med et lysosom, en vakuole som inneholder et sett med hydrolytiske enzymer.
Endocytose er formelt delt inn i pinocytose og fagocytose.
Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter
karakteristisk for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager). I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen
fagocytiske celler. Under fagocytose dannes store endocytiske vakuoler - fagosomer, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocytose
(pinocytose og fagocytose), såkalt fordi det går som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel,
partikler av sot eller fargestoffer.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, bretter, skiller små flytende medium volumer.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra cellemembranen.
overflaten og strekker seg dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Dette er de såkalte kantgropene. De kalles det fordi
På sidene av cytoplasmaet er plasmamembranen dekket, kledd, med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner grenser til og dekker små fremspring og groper. Disse hullene er
i nesten alle dyreceller opptar de omtrent 2 % av celleoverflaten. Grenselaget består hovedsakelig av klatrinproteinet assosiert med en rekke ekstra proteiner.
Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag langs omkretsen av det fremvoksende pinosomet.
Etter at den avgrensede vesikkelen skiller seg fra plasmolemmaet og begynner å bevege seg dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, og endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Reseptormediert endocytose. Effektiviteten av endocytose øker betydelig hvis den formidles av membranreseptorer som binder seg til molekyler av det absorberte stoffet eller molekyler som ligger på overflaten av det fagocyterte objektet - ligander (fra latin u^age - for å binde). Senere (etter absorpsjon av stoffet) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet. Et eksempel på en reseptormediert interaksjon er fagocytose av en bakteriell leukocytt.
Transcytose(fra lat. 1gash - gjennom, gjennom og gresk suYuz - celle) en prosess som er karakteristisk for noen typer celler, som kombinerer tegn på endocytose og eksocytose. En endocytisk vesikkel dannes på den ene celleoverflaten, som overføres til den motsatte celleoverflaten og som blir en eksocytisk vesikkel, frigjør innholdet til det ekstracellulære rommet.
Eksocytose
Plasmamembranen tar del i fjerning av stoffer fra cellen ved hjelp av eksocytose, en prosess som er det motsatte av endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster).
Makromolekyler som proteiner, nukleinsyrer, polysakkarider, lipoproteinkomplekser og andre passerer ikke gjennom cellemembraner, i motsetning til hvordan ioner og monomerer transporteres. Transporten av mikromolekyler, deres komplekser, partikler inn og ut av cellen skjer på en helt annen måte - gjennom vesikulær overføring. Dette begrepet betyr at ulike makromolekyler, biopolymerer eller deres komplekser ikke kan komme inn i cellen gjennom plasmamembranen. Og ikke bare gjennom det: noen cellemembraner er ikke i stand til transmembranoverføring av biopolymerer, med unntak av membraner som har spesielle proteinkompleksbærere - poriner (membraner av mitokondrier, plastider, peroksisomer). Makromolekyler kommer inn i cellen eller fra ett membranrom til et annet innelukket i vakuoler eller vesikler. Slik vesikulær overføring kan deles inn i to typer: eksocytose- fjerning av makromolekylære produkter fra cellen, og endocytose- absorpsjon av makromolekyler av cellen (fig. 133).
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. I en slik primærvakuole, eller i endosom, kan alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler komme inn, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet ved transmembranoverføring. Den viktigste biologiske betydningen av endocytose er oppkjøpet av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med lysosomet, en vakuole som inneholder et sett med hydrolytiske enzymer (se nedenfor).
Endocytose er formelt delt inn i pinocytose og fagocytose(Fig. 134). Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter, er karakteristiske for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager). Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocyto h (pinocytose og fagocytose), såkalt fordi det fortsetter som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel sotpartikler eller fargestoffer.
Uspesifikk endocytose er ofte ledsaget av innledende sorpsjon av innfangningsmaterialet av plasmamembranens glykokalyx. Glykokalyxen, på grunn av de sure gruppene i polysakkaridene, har en negativ ladning og binder seg godt til forskjellige positivt ladede grupper av proteiner. Med slik adsorpsjon absorberes uspesifikk endocytose, makromolekyler og små partikler (sure proteiner, ferritin, antistoffer, virioner, kolloidale partikler). Væskefase pinocytose fører til absorpsjon sammen med det flytende mediet av løselige molekyler som ikke binder seg til plasmalemmaet.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, folder, og skiller små volumer av det flytende mediet (fig. 135, 136). Den første typen forekomst av en pinocytisk vesikkel, pinosomer, er typisk for celler i tarmepitel, endotel, for amøber, den andre - for fagocytter og fibroblaster. Disse prosessene er avhengige av tilførsel av energi: respirasjonshemmere blokkerer disse prosessene.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra celleoverflaten og går dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Disse er de såkalte avgrensede groper. De kalles det fordi plasmamembranen fra siden av cytoplasmaet er dekket, kledd med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner så å si grenser til, dekker små fremspring, groper (fig. 137). Nesten alle dyreceller har disse gropene; de opptar omtrent 2% av celleoverflaten. Omgivende lag består hovedsakelig av protein clathrin assosiert med en rekke ekstra proteiner. Tre molekyler av clathrin, sammen med tre molekyler av et lavmolekylært protein, danner strukturen til et triskelion, som ligner et tre-strålet hakekors (fig. 138). Clathrin triskelions på den indre overflaten av gropene i plasmamembranen danner et løst nettverk bestående av femkanter og sekskanter, vanligvis ligner en kurv. Klathrinlaget dekker hele omkretsen av de separerende primære endocytiske vakuolene, avgrenset av vesikler.
Clathrin tilhører en av de såkalte artene. "dressing" proteiner (COP - belagte proteiner). Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag rundt omkretsen av det fremvoksende pinosomet, den primære endosomale vesikkelen - den "kantede" vesikkelen. i separasjonen av det primære endosomet er også proteiner involvert - dynaminer, som polymeriserer rundt halsen på skillevesikkelen (fig. 139).
Etter at den kantede vesikkelen skiller seg fra plasmolemmaet og begynner å bli overført dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Det ble funnet at membranene i de avgrensede gropene inneholder relativt lite kolesterol, noe som kan bestemme reduksjonen i membranstivhet og bidra til dannelse av bobler. Den biologiske betydningen av utseendet til en clathrin "belegg" langs periferien av vesiklene kan være at den gir adhesjon av de avgrensede vesiklene til elementene i cytoskjelettet og deres påfølgende transport i cellen, og forhindrer dem i å smelte sammen med hverandre .
Intensiteten av væskefase uspesifikk pinocytose kan være svært høy. Så epitelcellen i tynntarmen danner opptil 1000 pinosomer per sekund, og makrofager danner omtrent 125 pinosomer per minutt. Størrelsen på pinosomer er liten, deres nedre grense er 60–130 nm, men deres overflod fører til det faktum at under endocytose blir plasmolemma raskt erstattet, som om "brukt" på dannelsen av mange små vakuoler. Så i makrofager erstattes hele plasmamembranen på 30 minutter, i fibroblaster - om to timer.
Den videre skjebnen til endosomer kan være annerledes, noen av dem kan gå tilbake til celleoverflaten og smelte sammen med den, men de fleste av dem går inn i prosessen med intracellulær fordøyelse. Primære endosomer inneholder for det meste fremmede molekyler fanget i det flytende mediet og inneholder ikke hydrolytiske enzymer. endosomer kan smelte sammen mens de øker i størrelse. De smelter deretter sammen med primære lysosomer (se nedenfor), som introduserer enzymer i endosomhulen som hydrolyserer forskjellige biopolymerer. Virkningen av disse lysosomale hydrolasene forårsaker intracellulær fordøyelse - nedbrytning av polymerer til monomerer.
Som allerede nevnt, under fagocytose og pinocytose, mister celler et stort område av plasmolemma (se makrofager), som imidlertid raskt gjenopprettes under membranresirkulering, på grunn av tilbakeføring av vakuoler og deres inkorporering i plasmolemma. Dette skyldes det faktum at små vesikler kan skilles fra endosomer eller vakuoler, samt fra lysosomer, som igjen smelter sammen med plasmamembranen. Med slik resirkulering oppstår en slags "shuttle"-overføring av membraner: plasmolemma - pinosome - vakuole - plasmolemma. Dette fører til restaurering av det opprinnelige området av plasmamembranen. Det ble funnet at med en slik retur, membranresirkulering, holdes alt absorbert materiale tilbake i det gjenværende endosomet.
Spesifikk eller reseptor-mediert endocytose har en rekke forskjeller fra uspesifikke. Hovedsaken er at molekyler absorberes som det er spesifikke reseptorer for på plasmamembranen som bare er assosiert med denne typen molekyler. Ofte kalles slike molekyler som binder seg til reseptorproteiner på overflaten av celler ligander.
Reseptormediert endocytose ble først beskrevet i akkumulering av proteiner i fugleoocytter. Proteiner av eggeplommegranulat, vitellogeniner, syntetiseres i ulike vev, men så kommer de inn i eggstokkene med blodstrømmen, hvor de binder seg til spesielle membranreseptorer av oocytter og kommer deretter inn i cellen ved hjelp av endocytose, hvor eggeplommegranuler avsettes.
Et annet eksempel på selektiv endocytose er transport av kolesterol inn i cellen. Dette lipidet syntetiseres i leveren og danner i kombinasjon med andre fosfolipider og et proteinmolekyl det såkalte. low-density lipoprotein (LDL), som skilles ut av leverceller og fraktes gjennom hele kroppen av sirkulasjonssystemet (fig. 140). Spesielle plasmamembranreseptorer diffust plassert på overflaten av forskjellige celler gjenkjenner proteinkomponenten i LDL og danner et spesifikt reseptor-ligandkompleks. Etter dette beveger et slikt kompleks seg til sonen med avgrensede groper og blir internalisert - omgitt av en membran og nedsenket dypt inn i cytoplasmaet. Det har blitt vist at mutante reseptorer kan binde LDL, men ikke akkumuleres i området med avgrensede groper. I tillegg til LDL-reseptorer er det funnet mer enn to dusin andre stoffer involvert i reseptorendocytose av forskjellige stoffer, som alle bruker samme internaliseringsvei gjennom de avgrensede gropene. Sannsynligvis er deres rolle i akkumulering av reseptorer: en og samme kantede grop kan samle rundt 1000 reseptorer av forskjellige klasser. I fibroblaster er imidlertid LDL-reseptorklynger lokalisert i sonen med avgrensede groper selv i fravær av en ligand i mediet.
Den videre skjebnen til den absorberte LDL-partikkelen er at den gjennomgår forfall i sammensetningen sekundært lysosom. Etter nedsenking i cytoplasmaet til en avgrenset vesikkel lastet med LDL, er det et raskt tap av clathrinlaget, membranvesikler begynner å smelte sammen med hverandre og danner et endosom - en vakuole som inneholder absorberte LDL-partikler som fortsatt er assosiert med reseptorer på membranoverflaten . Deretter oppstår dissosiasjonen av ligand-reseptorkomplekset, små vakuoler spaltes fra endosomet, hvis membraner inneholder frie reseptorer. Disse vesiklene resirkuleres, inkorporeres i plasmamembranen, og dermed går reseptorene tilbake til celleoverflaten. Skjebnen til LDL er at de etter fusjon med lysosomer hydrolyseres til fritt kolesterol, som kan inkorporeres i cellemembraner.
Endosomer kjennetegnes ved en lavere pH-verdi (pH 4-5), et surere miljø enn andre cellevakuoler. Dette skyldes tilstedeværelsen i deres membraner av protonpumpeproteiner som pumper inn hydrogenioner med samtidig forbruk av ATP (H + -avhengig ATPase). Det sure miljøet i endosomer spiller en kritisk rolle i dissosiasjonen av reseptorer og ligander. I tillegg er et surt miljø optimalt for aktivering av hydrolytiske enzymer i lysosomer, som aktiveres ved fusjon av lysosomer med endosomer og fører til dannelsen endolysosomer, hvor spaltningen av absorberte biopolymerer skjer.
I noen tilfeller er skjebnen til dissosierte ligander ikke relatert til lysosomal hydrolyse. Så i noen celler, etter binding av plasmolemma-reseptorer til visse proteiner, synker clathrin-belagte vakuoler inn i cytoplasmaet og overføres til et annet område av cellen, hvor de smelter sammen igjen med plasmamembranen, og de bundne proteinene dissosieres fra reseptorer. Slik foregår overføringen, transcytose, av enkelte proteiner gjennom veggen i endotelcellen fra blodplasmaet til det intercellulære miljøet (fig. 141). Et annet eksempel på transcytose er overføring av antistoffer. Så hos pattedyr kan antistoffene til moren overføres til ungen gjennom melk. I dette tilfellet forblir reseptor-antistoffkomplekset uendret i endosomet.
Fagocytose
Som allerede nevnt er fagocytose en variant av endocytose og er assosiert med cellens absorpsjon av store aggregater av makromolekyler opp til levende eller døde celler. I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen til fagocytiske celler. Under fagocytose dannes det store endocytiske vakuoler - fagosomet, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
På overflaten av celler som er i stand til fagocytose (hos pattedyr er disse nøytrofiler og makrofager), er det et sett med reseptorer som interagerer med ligandproteiner. Ved bakterielle infeksjoner binder antistoffer mot bakterielle proteiner seg til overflaten av bakterieceller, og danner et lag der Fc-områdene til antistoffene ser utover. Dette laget gjenkjennes av spesifikke reseptorer på overflaten av makrofager og nøytrofiler, og ved bindingsstedene begynner absorpsjonen av bakterien ved å omslutte den med plasmamembranen til cellen (fig. 142).
Eksocytose
Plasmamembranen er involvert i fjerning av stoffer fra cellen ved hjelp av eksocytose- den omvendte prosessen med endocytose (se fig. 133).
Ved eksocytose nærmer intracellulære produkter innelukket i vakuoler eller vesikler og separert fra hyaloplasmaet med en membran plasmamembranen. Ved deres kontaktpunkter smelter plasmamembranen og vakuolmembranen sammen, og boblen tømmes ut i miljøet. Ved hjelp av eksocytose oppstår prosessen med resirkulering av membraner involvert i endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster). På samme måte fjernes noen polysakkarider (hemicelluloser) som er involvert i dannelsen av cellevegger fra cytoplasmaet til planteceller.
De fleste utskilte stoffene brukes av andre celler i flercellede organismer (utskillelse av melk, fordøyelsessaft, hormoner, etc.). Men ofte skiller celler ut stoffer til egne behov. For eksempel utføres veksten av plasmamembranen ved å legge inn deler av membranen som en del av eksocytiske vakuoler, noen av elementene i glykokalyxen utskilles av cellen i form av glykoproteinmolekyler, etc.
Hydrolytiske enzymer isolert fra celler ved eksocytose kan sorberes i glykokalyxlaget og gi membranbundet ekstracellulær spaltning av ulike biopolymerer og organiske molekyler. Membran ikke-cellulær fordøyelse er av stor betydning for dyr. Det ble funnet at i tarmepitelet til pattedyr i området med den såkalte børstegrensen til det absorberende epitelet, som er spesielt rikt på glykokalyx, finnes det en enorm mengde forskjellige enzymer. Noen av disse enzymene er av bukspyttkjertelopprinnelse (amylase, lipaser, ulike proteinaser, etc.), og noen skilles ut av selve epitelcellene (eksohydrolaser, som bryter ned hovedsakelig oligomerer og dimerer med dannelse av transporterte produkter).
©2015-2019 nettsted
Alle rettigheter tilhører deres forfattere. Dette nettstedet krever ikke forfatterskap, men tilbyr gratis bruk.
Opprettelsesdato for side: 2016-04-15
Store molekyler av biopolymerer transporteres praktisk talt ikke gjennom membraner, og likevel kan de komme inn i cellen som følge av endocytose. Den er delt inn i fagocytose og pinocytose. Disse prosessene er assosiert med kraftig aktivitet og mobilitet av cytoplasma. Fagocytose er fangst og absorpsjon av store partikler av en celle (noen ganger til og med hele celler og deres deler). Fagocytose og pinocytose forløper veldig likt, derfor gjenspeiler disse konseptene bare forskjellen i volumene av absorberte stoffer. Felles for dem er at de absorberte stoffene på celleoverflaten er omgitt av en membran i form av en vakuole, som beveger seg inne i cellen (enten en fagocytisk eller pinocytisk vesikkel. Disse prosessene er forbundet med energiforbruk; opphør av ATP-syntese hemmer dem fullstendig. for eksempel veggene i tarmene, mange mikrovilli, betydelig øke overflaten som absorpsjon skjer gjennom. Plasmamembranen er også involvert i fjerning av stoffer fra cellen, dette skjer i prosessen eksocytose. Slik skilles hormoner, polysakkarider, proteiner, fettdråper og andre celleprodukter ut. De er innelukket i membranbundne vesikler og nærmer seg plasmalemmaet. Begge membranene smelter sammen og innholdet i vesikkelen slippes ut i miljøet rundt cellen.
Celler er også i stand til å absorbere makromolekyler og partikler ved hjelp av en lignende eksocytose mekanisme, men i omvendt rekkefølge. Absorbert materiale er gradvis omgitt av et lite område plasmamembran, som først invaginerer og deretter deler seg av og dannes intracellulær vesikkel som inneholder materiale fanget av cellen. Denne prosessen med dannelse av intracellulære vesikler rundt materialet som absorberes av cellen kalles endocytose.
Avhengig av størrelsen på vesiklene som dannes, skilles to typer endocytose:
1) pinocytose- absorpsjon av væske og oppløste stoffer gjennom små bobler, og
2) fagocytose- absorpsjon av store partikler som mikroorganismer eller celleavfall. I dette tilfellet dannes det store bobler, kalt vakuoler og absorpsjon av korpuskulært materiale: bakterier, store virus, døende egne celler i kroppen eller fremmede celler, som for eksempel erytrocytter av forskjellige typer, utføres av celler ( makrofager ,nøytrofiler)
Væske og oppløste stoffer tas kontinuerlig opp av de fleste celler gjennom pinocytose, mens store partikler hovedsakelig tas opp av spesialiserte celler - fagocytter. Derfor brukes begrepene "pinocytose" og "endocytose" vanligvis i samme betydning.
pinocytose karakterisert ved absorpsjon og intracellulær ødeleggelse av makromolekylære forbindelser, slik som proteiner og proteinkomplekser, nukleinsyrer, polysakkarider, lipoproteiner. Gjenstanden for pinocytose som en faktor for uspesifikk immunforsvar er spesielt giftstoffene til mikroorganismer. Adhesjon av stoffer på celleoverflaten fører til lokal invaginasjon (invaginasjon) av membranen, som kulminerer med dannelsen av en pinocytisk vesikkel av svært liten størrelse (ca. 0,1 mikron). Flere sammenslåtte bobler danner en større formasjon - pinosom. I neste trinn smelter pinosomene sammen med lysosomer inneholder hydrolytiske enzymer som bryter ned polymermolekyler til monomerer. I tilfeller der prosessen med pinocytose realiseres gjennom reseptorapparatet, i pinosomer, før fusjon med lysosomer, observeres løsrivelsen av fangede molekyler fra reseptorer, som, som en del av dattervesiklene, går tilbake til celleoverflaten.
vesikulær overføring eksocytose endocytose
endosom
pinocytose og fagocytose(Fig. 134). karakteristisk for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager).
Uspesifikk endocyto fra partikler av sot eller fargestoffer.
overflaten og strekker seg dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Disse er de såkalte avgrensede groper clathrin
Spesifikk eller reseptor-mediert ligander.
sekundært lysosom
endolysosomer
Fagocytose
fagosomet fagolysosomer.
Eksocytose
eksocytose
©2015-2019 nettsted
Alle rettigheter tilhører deres forfattere. Dette nettstedet krever ikke forfatterskap, men tilbyr gratis bruk.
Opprettelsesdato for side: 2016-04-15
Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter
Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
(pinocytose og fagocytose), såkalt fordi det går som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel,
partikler av sot eller fargestoffer.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra cellemembranen.
Transcytose
Eksocytose
Ved eksocytose nærmer intracellulære produkter innelukket i vakuoler eller vesikler og separert fra hyaloplasmaet med en membran plasmamembranen. Ved deres kontaktpunkter smelter plasmamembranen og vakuolmembranen sammen, og boblen tømmes ut i miljøet. Ved hjelp av eksocytose oppstår prosessen med resirkulering av membraner involvert i endocytose.
Vesikulær overføring kan deles inn i to typer: eksocytose - fjerning av makromolekylære produkter fra cellen, og endocytose - absorpsjon av makromolekyler av cellen.
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. Alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler kan gå inn i en slik primær vakuole, eller endosom, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet gjennom transmembranoverføring.
Den viktigste biologiske betydningen av endocytose er anskaffelsen av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med et lysosom, en vakuole som inneholder et sett med hydrolytiske enzymer.
Endocytose er formelt delt inn i pinocytose og fagocytose.
Fagocytose - fangst og absorpsjon av store partikler av en celle (noen ganger til og med celler eller deres deler) - ble først beskrevet av I.I. Mechnikov. Fagocytose, evnen til å fange store partikler av en celle, finnes blant dyreceller, både encellede (for eksempel amøber, noen rov-ciliater) og spesialiserte celler fra flercellede dyr. Spesialiserte celler, fagocytter
karakteristisk for både virvelløse dyr (amoebocytter av blod eller hulromsvæske) og vertebrater (nøytrofiler og makrofager). I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen
fagocytiske celler. Under fagocytose dannes store endocytiske vakuoler - fagosomer, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
Pinocytose ble opprinnelig definert som absorpsjon av vann eller vandige løsninger av ulike stoffer av cellen. Det er nå kjent at både fagocytose og pinocytose forløper veldig likt, og derfor kan bruken av disse begrepene bare gjenspeile forskjeller i volumer og masse av absorberte stoffer. Felles for disse prosessene er at de absorberte stoffene på overflaten av plasmamembranen er omgitt av en membran i form av en vakuole – et endosom, som beveger seg inne i cellen.
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocytose
(pinocytose og fagocytose), såkalt fordi det går som automatisk og ofte kan føre til fangst og absorpsjon av stoffer som er helt fremmede eller likegyldige for cellen, for eksempel,
partikler av sot eller fargestoffer.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, bretter, skiller små flytende medium volumer.
Etter denne omorganiseringen av overflaten følger prosessen med adhesjon og fusjon av kontaktmembranene, noe som fører til dannelsen av en penicytisk vesikkel (pinosom), som løsner fra cellemembranen.
overflaten og strekker seg dypt inn i cytoplasmaet. Både uspesifikk og reseptorendocytose, som fører til spaltning av membranvesikler, forekommer i spesialiserte områder av plasmamembranen. Dette er de såkalte kantgropene. De kalles det fordi
På sidene av cytoplasmaet er plasmamembranen dekket, kledd, med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner grenser til og dekker små fremspring og groper. Disse hullene er
i nesten alle dyreceller opptar de omtrent 2 % av celleoverflaten. Grenselaget består hovedsakelig av klatrinproteinet assosiert med en rekke ekstra proteiner.
Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag langs omkretsen av det fremvoksende pinosomet.
Etter at den avgrensede vesikkelen skiller seg fra plasmolemmaet og begynner å bevege seg dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, og endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Reseptormediert endocytose. Effektiviteten av endocytose øker betydelig hvis den formidles av membranreseptorer som binder seg til molekyler av det absorberte stoffet eller molekyler som ligger på overflaten av det fagocyterte objektet - ligander (fra latin u^age - for å binde). Senere (etter absorpsjon av stoffet) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet. Et eksempel på en reseptormediert interaksjon er fagocytose av en bakteriell leukocytt.
Transcytose(fra lat. 1gash - gjennom, gjennom og gresk suYuz - celle) en prosess som er karakteristisk for noen typer celler, som kombinerer tegn på endocytose og eksocytose. En endocytisk vesikkel dannes på den ene celleoverflaten, som overføres til den motsatte celleoverflaten og som blir en eksocytisk vesikkel, frigjør innholdet til det ekstracellulære rommet.
Eksocytose
Plasmamembranen tar del i fjerning av stoffer fra cellen ved hjelp av eksocytose, en prosess som er det motsatte av endocytose.
Ved eksocytose nærmer intracellulære produkter innelukket i vakuoler eller vesikler og separert fra hyaloplasmaet med en membran plasmamembranen. Ved deres kontaktpunkter smelter plasmamembranen og vakuolmembranen sammen, og boblen tømmes ut i miljøet. Ved hjelp av eksocytose oppstår prosessen med resirkulering av membraner involvert i endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster).
41 .Endoplasmatisk retikulum (retikulum).
I et lysmikroskop i fibriblaster etter fiksering og farging kan man se at periferien av cellene (ektoplasma) farges svakt, mens den sentrale delen av cellene (endoplasma) oppfatter fargestoffer godt. Så K. Porter i 1945 så i et elektronmikroskop at den endoplasmatiske sonen er fylt med et stort antall små vakuoler og kanaler som forbinder seg med hverandre og danner noe som et løst nettverk (retikulum). Det ble sett at stablene av disse vakuolene og tubuli var begrenset av tynne membraner. Så det ble oppdaget endoplasmatisk retikulum, eller endoplasmatisk retikulum. Senere, på 1950-tallet, ved hjelp av metoden med ultratynne seksjoner, var det mulig å belyse strukturen til denne formasjonen og å oppdage dens heterogenitet. Det viktigste viste seg å være at endoplasmatisk retikulum (ER) finnes i nesten alle eukaryoter.
En slik elektronmikroskopisk analyse gjorde det mulig å skille to typer ER: granulær (grov) og glatt.
Del 3. Transmembranbevegelse av makromolekyler
Makromolekyler kan transporteres over plasmamembranen. Prosessen der celler tar inn store molekyler kalles endocytose. Noen av disse molekylene (for eksempel polysakkarider, proteiner og polynukleotider) tjener som en kilde til næringsstoffer. Endocytose gjør det også mulig å regulere innholdet av visse membrankomponenter, spesielt hormonreseptorer. Endocytose kan brukes til å studere cellulære funksjoner mer detaljert. Celler av en type kan transformeres med en annen type DNA og dermed endre funksjon eller fenotype.
I slike eksperimenter brukes ofte spesifikke gener, noe som gir en unik mulighet til å studere mekanismene for deres regulering. Transformasjonen av celler ved hjelp av DNA utføres ved endocytose - dette er måten DNA kommer inn i cellen. Transformasjon utføres vanligvis i nærvær av kalsiumfosfat, siden Ca 2+ stimulerer endocytose og DNA-utfelling, noe som letter dens inntreden i cellen ved endocytose.
Makromolekyler forlater cellen eksocytose. Ved både endocytose og eksocytose dannes det vesikler som smelter sammen med plasmamembranen eller løsner fra den.
3.1. Endocytose: typer endocytose og mekanisme
Alle eukaryote celler en del av plasmamembranen er hele tiden inne i cytoplasmaet. Dette skjer som et resultat invaginasjon av et fragment av plasmamembranen, utdanning endocytisk vesikkel , lukke halsen på vesikkelen og snøre den inn i cytoplasmaet sammen med innholdet (Fig. 18). Deretter kan vesikler smelte sammen med andre membranstrukturer og dermed overføre innholdet til andre cellulære rom eller til og med tilbake til det ekstracellulære rommet. De fleste endocytiske vesikler smelter sammen med primære lysosomer og danner sekundære lysosomer, som inneholder hydrolytiske enzymer og er spesialiserte organeller. Makromolekyler fordøyes i dem til aminosyrer, enkle sukkerarter og nukleotider, som diffunderer fra vesiklene og utnyttes i cytoplasmaet.
For endocytose trenger du:
1) energi, kilden som vanligvis er ATP;
2) ekstracellulært Ca 2+;
3) kontraktile elementer i cellen(sannsynligvis mikrofilamentsystemer).
Endocytose kan deles inn tre hovedtyper:
1. Fagocytose kun utført involverer spesialiserte celler (Fig. 19), slik som makrofager og granulocytter. Under fagocytose absorberes store partikler - virus, bakterier, celler eller deres fragmenter. Makrofager er eksepsjonelt aktive i denne forbindelse og kan skru på et volum på 25 % av sitt eget volum på 1 time. Dette internaliserer 3 % av plasmamembranen hvert minutt, eller hele membranen hvert 30. minutt.
 2. pinocytose finnes i alle celler. Med den, cellen absorberer væsker
og komponenter oppløst i den (fig. 20). Flytende fase pinocytose er ikke-selektiv prosess
, hvor mengden av det oppløste stoffet absorbert i sammensetningen av vesiklene ganske enkelt er proporsjonal med konsentrasjonen i den ekstracellulære væsken. Slike vesikler dannes utelukkende aktivt. For eksempel, i fibroblaster, er internaliseringshastigheten av plasmamembranen 1/3 av hastigheten som er karakteristisk for makrofager. I dette tilfellet forbrukes membranen raskere enn den syntetiseres. Samtidig endres ikke cellens overflate og volum mye, noe som indikerer at membranen gjenopprettes på grunn av eksocytose eller på grunn av dens re-inklusjon i samme hastighet som den konsumeres.
2. pinocytose finnes i alle celler. Med den, cellen absorberer væsker
og komponenter oppløst i den (fig. 20). Flytende fase pinocytose er ikke-selektiv prosess
, hvor mengden av det oppløste stoffet absorbert i sammensetningen av vesiklene ganske enkelt er proporsjonal med konsentrasjonen i den ekstracellulære væsken. Slike vesikler dannes utelukkende aktivt. For eksempel, i fibroblaster, er internaliseringshastigheten av plasmamembranen 1/3 av hastigheten som er karakteristisk for makrofager. I dette tilfellet forbrukes membranen raskere enn den syntetiseres. Samtidig endres ikke cellens overflate og volum mye, noe som indikerer at membranen gjenopprettes på grunn av eksocytose eller på grunn av dens re-inklusjon i samme hastighet som den konsumeres.
 3. Reseptormediert endocytose(reopptak av nevrotransmitter) - endocytose, der membranreseptorer binder seg til molekyler av det absorberte stoffet, eller molekyler lokalisert på overflaten av det fagocyterte objektet - ligander (fra lat. ligare–binde(Fig. 21) )
. Senere (etter absorpsjon av et stoff eller objekt) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet.
3. Reseptormediert endocytose(reopptak av nevrotransmitter) - endocytose, der membranreseptorer binder seg til molekyler av det absorberte stoffet, eller molekyler lokalisert på overflaten av det fagocyterte objektet - ligander (fra lat. ligare–binde(Fig. 21) )
. Senere (etter absorpsjon av et stoff eller objekt) spaltes reseptor-ligandkomplekset, og reseptorene kan igjen gå tilbake til plasmalemmaet.
Et eksempel på reseptormediert endocytose er fagocytose av en bakterie med en leukocytt. Siden leukocyttens plasmolemma har reseptorer for immunglobuliner (antistoffer), øker fagocytosehastigheten hvis overflaten av bakteriecelleveggen er dekket med antistoffer (opsoniner - fra det greske opson–krydder).
 Reseptormediert endocytose er en aktiv spesifikk prosess der cellemembranen buler inn i cellen og danner avgrensede groper
. Den intracellulære siden av den avgrensede gropen inneholder sett med adaptive proteiner
(adaptin, clathrin, som bestemmer den nødvendige krumningen av bulen, og andre proteiner) (Fig. 22). Når liganden er bundet fra miljøet som omgir cellen, danner de avgrensede gropene intracellulære vesikler (kantede vesikler). Reseptormediert endocytose slås på for raskt og kontrollert opptak av cellen av den aktuelle liganden. Disse vesiklene mister raskt grensen og smelter sammen med hverandre, og danner større vesikler - endosomer.
Reseptormediert endocytose er en aktiv spesifikk prosess der cellemembranen buler inn i cellen og danner avgrensede groper
. Den intracellulære siden av den avgrensede gropen inneholder sett med adaptive proteiner
(adaptin, clathrin, som bestemmer den nødvendige krumningen av bulen, og andre proteiner) (Fig. 22). Når liganden er bundet fra miljøet som omgir cellen, danner de avgrensede gropene intracellulære vesikler (kantede vesikler). Reseptormediert endocytose slås på for raskt og kontrollert opptak av cellen av den aktuelle liganden. Disse vesiklene mister raskt grensen og smelter sammen med hverandre, og danner større vesikler - endosomer.
clathrin- intracellulært protein, hovedkomponenten i membranen til avgrensede vesikler dannet under reseptorendocytose (fig. 23).
Tre clathrin-molekyler er assosiert med hverandre i den C-terminale enden på en slik måte at clathrin-trimeren har form som en triskelion. Som et resultat av polymerisering danner clathrin et lukket tredimensjonalt nettverk som ligner en fotball. Størrelsen på clathrin vesikler er omtrent 100 nm.
Avgrensede groper kan okkupere opptil 2% av overflaten til noen celler. Endocytiske vesikler som inneholder lipoproteiner med lav tetthet (LDL) og deres reseptorer smelter sammen med lysosomer i cellen. Reseptorene frigjøres og returneres til overflaten av cellemembranen, og LDL-apoproteinet spaltes og den tilsvarende kolesterolesteren metaboliseres. Syntese av LDL-reseptorer reguleres av sekundære eller tertiære produkter av pinocytose, dvs. stoffer som dannes under metabolismen av LDL, slik som kolesterol.
3.2. Eksocytose: kalsiumavhengig og kalsiumuavhengig.
De fleste celler frigjør makromolekyler til miljøet ved eksocytose . Denne prosessen spiller også en rolle i membranfornyelse når dets komponenter syntetisert i Golgi-apparatet leveres som en del av vesikler til plasmamembranen (fig. 24).
 |
Ris. 24. Sammenligning av mekanismene for endocytose og eksocytose.
Mellom ekso- og endocytose, i tillegg til forskjellen i bevegelsesretningen til stoffer, er det en annen betydelig forskjell: når eksocytose fortsette fusjon av to indre cytoplasmatiske monolag , mens kl endocyose de ytre monolagene smelter sammen.
Stoffer frigjort ved eksocytose, kan deles i tre kategorier:
1) stoffer som binder seg til celleoverflaten og å bli perifere proteiner, slik som antigener;
2) stoffer som inngår i den ekstracellulære matrisen f.eks. kollagen og glykosaminoglykaner;
3) stoffer som slippes ut i det ekstracellulære miljøet og fungerer som signalmolekyler for andre celler.
Eukaryoter er utmerkede to typer eksocytose:
1. Kalsiumuavhengig konstitutiv eksocytose forekommer i nesten alle eukaryote celler. Det er en nødvendig prosess å bygge en ekstracellulær matrise og levere proteiner til den ytre cellemembranen. I dette tilfellet blir sekretoriske vesikler levert til celleoverflaten og smelter sammen med den ytre membranen når de dannes.
2. kalsiumavhengig ikke-konstitutiv eksocytose forekommer, for eksempel, i kjemiske synapser eller celler som produserer makromolekylære hormoner. Denne eksocytosen tjener f.eks. å isolere nevrotransmittere. Ved denne typen eksocytose akkumuleres sekretoriske vesikler i cellen, og prosessen med utgivelsen utløses av et visst signal mediert av en rask økning i konsentrasjonen kalsiumioner i cytosolen til cellen. I presynaptiske membraner utføres prosessen av et spesielt kalsiumavhengig proteinkompleks SNARE.
Vesikulær transport: endocytose og eksocytose
Makromolekyler som proteiner, nukleinsyrer, polysakkarider, lipoproteinkomplekser og andre passerer ikke gjennom cellemembraner, i motsetning til hvordan ioner og monomerer transporteres. Transporten av mikromolekyler, deres komplekser, partikler inn og ut av cellen skjer på en helt annen måte - gjennom vesikulær overføring. Dette begrepet betyr at ulike makromolekyler, biopolymerer eller deres komplekser ikke kan komme inn i cellen gjennom plasmamembranen. Og ikke bare gjennom det: noen cellemembraner er ikke i stand til transmembranoverføring av biopolymerer, med unntak av membraner som har spesielle proteinkompleksbærere - poriner (membraner av mitokondrier, plastider, peroksisomer). Makromolekyler kommer inn i cellen eller fra ett membranrom til et annet innelukket i vakuoler eller vesikler. Slik vesikulær overføring kan deles inn i to typer: eksocytose- fjerning av makromolekylære produkter fra cellen, og endocytose- absorpsjon av makromolekyler av cellen (fig. 133).
Under endocytose fanger en viss del av plasmalemmaet, så å si, omslutter det ekstracellulære materialet, og omslutter det i en membranvakuole som har oppstått på grunn av invaginasjonen av plasmamembranen. I en slik primærvakuole, eller i endosom, kan alle biopolymerer, makromolekylære komplekser, deler av celler eller til og med hele celler komme inn, hvor de deretter brytes ned, depolymeriseres til monomerer, som kommer inn i hyaloplasmaet ved transmembranoverføring. Den viktigste biologiske betydningen av endocytose er oppkjøpet av byggesteiner gjennom intracellulær fordøyelse, som utføres i det andre stadiet av endocytose etter fusjonen av det primære endosomet med lysosomet, en vakuole som inneholder et sett med hydrolytiske enzymer (se nedenfor).
Endocytose er formelt delt inn i pinocytose og fagocytose
Endocytose, inkludert pinocytose og fagocytose, kan være uspesifikk eller konstitutiv, permanent og spesifikk, mediert av reseptorer (reseptor). Uspesifikk endocyto
Uspesifikk endocytose er ofte ledsaget av innledende sorpsjon av innfangningsmaterialet av plasmamembranens glykokalyx. Glykokalyxen, på grunn av de sure gruppene i polysakkaridene, har en negativ ladning og binder seg godt til forskjellige positivt ladede grupper av proteiner. Med slik adsorpsjon absorberes uspesifikk endocytose, makromolekyler og små partikler (sure proteiner, ferritin, antistoffer, virioner, kolloidale partikler). Væskefase pinocytose fører til absorpsjon sammen med det flytende mediet av løselige molekyler som ikke binder seg til plasmalemmaet.
På neste stadium skjer en endring i morfologien til celleoverflaten: det er enten utseendet til små invaginasjoner av plasmamembranen, invaginasjon, eller det er utseendet på celleoverflaten av utvekster, folder eller "frills" (rafl - på engelsk), som så å si overlapper, folder, og skiller små volumer av det flytende mediet (fig. 135, 136). Den første typen forekomst av en pinocytisk vesikkel, pinosomer, er typisk for celler i tarmepitel, endotel, for amøber, den andre - for fagocytter og fibroblaster. Disse prosessene er avhengige av tilførsel av energi: respirasjonshemmere blokkerer disse prosessene.
avgrensede groper. De kalles det fordi plasmamembranen fra siden av cytoplasmaet er dekket, kledd med et tynt (ca. 20 nm) fibrøst lag, som på ultratynne seksjoner så å si grenser til, dekker små fremspring, groper (fig. 137). Nesten alle dyreceller har disse gropene; de opptar omtrent 2% av celleoverflaten. Omgivende lag består hovedsakelig av protein clathrin assosiert med en rekke ekstra proteiner. Tre molekyler av clathrin, sammen med tre molekyler av et lavmolekylært protein, danner strukturen til et triskelion, som ligner et tre-strålet hakekors (fig. 138). Clathrin triskelions på den indre overflaten av gropene i plasmamembranen danner et løst nettverk bestående av femkanter og sekskanter, vanligvis ligner en kurv. Klathrinlaget dekker hele omkretsen av de separerende primære endocytiske vakuolene, avgrenset av vesikler.
Clathrin tilhører en av de såkalte artene. "dressing" proteiner (COP - belagte proteiner). Disse proteinene binder seg til integrerte reseptorproteiner fra siden av cytoplasmaet og danner et forbindingslag rundt omkretsen av det fremvoksende pinosomet, den primære endosomale vesikkelen - den "kantede" vesikkelen. i separasjonen av det primære endosomet er også proteiner involvert - dynaminer, som polymeriserer rundt halsen på skillevesikkelen (fig. 139).
Etter at den kantede vesikkelen skiller seg fra plasmolemmaet og begynner å bli overført dypt inn i cytoplasmaet, desintegrerer clathrinlaget, dissosieres, endosommembranen (pinosomer) får sin vanlige form. Etter tapet av clathrinlaget begynner endosomene å smelte sammen.
Det ble funnet at membranene i de avgrensede gropene inneholder relativt lite kolesterol, noe som kan bestemme reduksjonen i membranstivhet og bidra til dannelse av bobler. Den biologiske betydningen av utseendet til en clathrin "belegg" langs periferien av vesiklene kan være at den gir adhesjon av de avgrensede vesiklene til elementene i cytoskjelettet og deres påfølgende transport i cellen, og forhindrer dem i å smelte sammen med hverandre .
Intensiteten av væskefase uspesifikk pinocytose kan være svært høy. Så epitelcellen i tynntarmen danner opptil 1000 pinosomer per sekund, og makrofager danner omtrent 125 pinosomer per minutt. Størrelsen på pinosomer er liten, deres nedre grense er 60–130 nm, men deres overflod fører til det faktum at under endocytose blir plasmolemma raskt erstattet, som om "brukt" på dannelsen av mange små vakuoler. Så i makrofager erstattes hele plasmamembranen på 30 minutter, i fibroblaster - om to timer.
Den videre skjebnen til endosomer kan være annerledes, noen av dem kan gå tilbake til celleoverflaten og smelte sammen med den, men de fleste av dem går inn i prosessen med intracellulær fordøyelse. Primære endosomer inneholder for det meste fremmede molekyler fanget i det flytende mediet og inneholder ikke hydrolytiske enzymer. endosomer kan smelte sammen mens de øker i størrelse. De smelter deretter sammen med primære lysosomer (se nedenfor), som introduserer enzymer i endosomhulen som hydrolyserer forskjellige biopolymerer. Virkningen av disse lysosomale hydrolasene forårsaker intracellulær fordøyelse - nedbrytning av polymerer til monomerer.
Som allerede nevnt, under fagocytose og pinocytose, mister celler et stort område av plasmolemma (se makrofager), som imidlertid raskt gjenopprettes under membranresirkulering, på grunn av tilbakeføring av vakuoler og deres inkorporering i plasmolemma. Dette skyldes det faktum at små vesikler kan skilles fra endosomer eller vakuoler, samt fra lysosomer, som igjen smelter sammen med plasmamembranen. Med slik resirkulering oppstår en slags "shuttle"-overføring av membraner: plasmolemma - pinosome - vakuole - plasmolemma. Dette fører til restaurering av det opprinnelige området av plasmamembranen. Det ble funnet at med en slik retur, membranresirkulering, holdes alt absorbert materiale tilbake i det gjenværende endosomet.
Spesifikk eller reseptor-mediert endocytose har en rekke forskjeller fra uspesifikke. Hovedsaken er at molekyler absorberes som det er spesifikke reseptorer for på plasmamembranen som bare er assosiert med denne typen molekyler. Ofte kalles slike molekyler som binder seg til reseptorproteiner på overflaten av celler ligander.
Reseptormediert endocytose ble først beskrevet i akkumulering av proteiner i fugleoocytter. Proteiner av eggeplommegranulat, vitellogeniner, syntetiseres i ulike vev, men så kommer de inn i eggstokkene med blodstrømmen, hvor de binder seg til spesielle membranreseptorer av oocytter og kommer deretter inn i cellen ved hjelp av endocytose, hvor eggeplommegranuler avsettes.
Et annet eksempel på selektiv endocytose er transport av kolesterol inn i cellen. Dette lipidet syntetiseres i leveren og danner i kombinasjon med andre fosfolipider og et proteinmolekyl det såkalte. low-density lipoprotein (LDL), som skilles ut av leverceller og fraktes gjennom hele kroppen av sirkulasjonssystemet (fig. 140). Spesielle plasmamembranreseptorer diffust plassert på overflaten av forskjellige celler gjenkjenner proteinkomponenten i LDL og danner et spesifikt reseptor-ligandkompleks. Etter dette beveger et slikt kompleks seg til sonen med avgrensede groper og blir internalisert - omgitt av en membran og nedsenket dypt inn i cytoplasmaet. Det har blitt vist at mutante reseptorer kan binde LDL, men ikke akkumuleres i området med avgrensede groper. I tillegg til LDL-reseptorer er det funnet mer enn to dusin andre stoffer involvert i reseptorendocytose av forskjellige stoffer, som alle bruker samme internaliseringsvei gjennom de avgrensede gropene. Sannsynligvis er deres rolle i akkumulering av reseptorer: en og samme kantede grop kan samle rundt 1000 reseptorer av forskjellige klasser. I fibroblaster er imidlertid LDL-reseptorklynger lokalisert i sonen med avgrensede groper selv i fravær av en ligand i mediet.
Den videre skjebnen til den absorberte LDL-partikkelen er at den gjennomgår forfall i sammensetningen sekundært lysosom. Etter nedsenking i cytoplasmaet til en avgrenset vesikkel lastet med LDL, er det et raskt tap av clathrinlaget, membranvesikler begynner å smelte sammen med hverandre og danner et endosom - en vakuole som inneholder absorberte LDL-partikler som fortsatt er assosiert med reseptorer på membranoverflaten . Deretter oppstår dissosiasjonen av ligand-reseptorkomplekset, små vakuoler spaltes fra endosomet, hvis membraner inneholder frie reseptorer. Disse vesiklene resirkuleres, inkorporeres i plasmamembranen, og dermed går reseptorene tilbake til celleoverflaten. Skjebnen til LDL er at de etter fusjon med lysosomer hydrolyseres til fritt kolesterol, som kan inkorporeres i cellemembraner.
Endosomer kjennetegnes ved en lavere pH-verdi (pH 4-5), et surere miljø enn andre cellevakuoler. Dette skyldes tilstedeværelsen i deres membraner av protonpumpeproteiner som pumper inn hydrogenioner med samtidig forbruk av ATP (H + -avhengig ATPase). Det sure miljøet i endosomer spiller en kritisk rolle i dissosiasjonen av reseptorer og ligander. I tillegg er et surt miljø optimalt for aktivering av hydrolytiske enzymer i lysosomer, som aktiveres ved fusjon av lysosomer med endosomer og fører til dannelsen endolysosomer, hvor spaltningen av absorberte biopolymerer skjer.
I noen tilfeller er skjebnen til dissosierte ligander ikke relatert til lysosomal hydrolyse. Så i noen celler, etter binding av plasmolemma-reseptorer til visse proteiner, synker clathrin-belagte vakuoler inn i cytoplasmaet og overføres til et annet område av cellen, hvor de smelter sammen igjen med plasmamembranen, og de bundne proteinene dissosieres fra reseptorer. Slik foregår overføringen, transcytose, av enkelte proteiner gjennom veggen i endotelcellen fra blodplasmaet til det intercellulære miljøet (fig. 141). Et annet eksempel på transcytose er overføring av antistoffer. Så hos pattedyr kan antistoffene til moren overføres til ungen gjennom melk. I dette tilfellet forblir reseptor-antistoffkomplekset uendret i endosomet.
Fagocytose
Som allerede nevnt er fagocytose en variant av endocytose og er assosiert med cellens absorpsjon av store aggregater av makromolekyler opp til levende eller døde celler. I tillegg til pinocytose kan fagocytose være uspesifikk (for eksempel absorpsjon av partikler av kolloidalt gull eller dekstranpolymer av fibroblaster eller makrofager) og spesifikk, mediert av reseptorer på overflaten av plasmamembranen til fagocytiske celler. Under fagocytose dannes det store endocytiske vakuoler - fagosomet, som deretter smelter sammen med lysosomer for å danne fagolysosomer.
På overflaten av celler som er i stand til fagocytose (hos pattedyr er disse nøytrofiler og makrofager), er det et sett med reseptorer som interagerer med ligandproteiner. Ved bakterielle infeksjoner binder antistoffer mot bakterielle proteiner seg til overflaten av bakterieceller, og danner et lag der Fc-områdene til antistoffene ser utover. Dette laget gjenkjennes av spesifikke reseptorer på overflaten av makrofager og nøytrofiler, og ved bindingsstedene begynner absorpsjonen av bakterien ved å omslutte den med plasmamembranen til cellen (fig. 142).
Eksocytose
Plasmamembranen er involvert i fjerning av stoffer fra cellen ved hjelp av eksocytose- den omvendte prosessen med endocytose (se fig. 133).
Ved eksocytose nærmer intracellulære produkter innelukket i vakuoler eller vesikler og separert fra hyaloplasmaet med en membran plasmamembranen. Ved deres kontaktpunkter smelter plasmamembranen og vakuolmembranen sammen, og boblen tømmes ut i miljøet. Ved hjelp av eksocytose oppstår prosessen med resirkulering av membraner involvert i endocytose.
Eksocytose er assosiert med frigjøring av forskjellige stoffer syntetisert i cellen. Celler som skiller ut, frigjør stoffer til miljøet, kan produsere og frigjøre forbindelser med lav molekylvekt (acetylkolin, biogene aminer, etc.), så vel som i de fleste tilfeller makromolekyler (peptider, proteiner, lipoproteiner, peptidoglykaner, etc.). Eksocytose eller sekresjon oppstår i de fleste tilfeller som respons på et eksternt signal (nerveimpuls, hormoner, mediatorer, etc.). Selv om det i noen tilfeller forekommer eksocytose konstant (utskillelse av fibronektin og kollagen av fibroblaster). På samme måte fjernes noen polysakkarider (hemicelluloser) som er involvert i dannelsen av cellevegger fra cytoplasmaet til planteceller.
De fleste utskilte stoffene brukes av andre celler i flercellede organismer (utskillelse av melk, fordøyelsessaft, hormoner, etc.). Men ofte skiller celler ut stoffer til egne behov. For eksempel utføres veksten av plasmamembranen ved å legge inn deler av membranen som en del av eksocytiske vakuoler, noen av elementene i glykokalyxen utskilles av cellen i form av glykoproteinmolekyler, etc.
Hydrolytiske enzymer isolert fra celler ved eksocytose kan sorberes i glykokalyxlaget og gi membranbundet ekstracellulær spaltning av ulike biopolymerer og organiske molekyler. Membran ikke-cellulær fordøyelse er av stor betydning for dyr. Det ble funnet at i tarmepitelet til pattedyr i området med den såkalte børstegrensen til det absorberende epitelet, som er spesielt rikt på glykokalyx, finnes det en enorm mengde forskjellige enzymer. Noen av disse enzymene er av bukspyttkjertelopprinnelse (amylase, lipaser, ulike proteinaser, etc.), og noen skilles ut av selve epitelcellene (eksohydrolaser, som bryter ned hovedsakelig oligomerer og dimerer med dannelse av transporterte produkter).
Reseptorrollen til plasmalemmaet
Vi har allerede møtt denne funksjonen til plasmamembranen når vi ble kjent med dens transportfunksjoner. Bærerproteiner og pumper er også reseptorer som gjenkjenner og samhandler med visse ioner. Reseptorproteiner binder seg til ligander og deltar i utvalget av molekyler som kommer inn i celler.
Membranproteiner eller glykokalykselementer - glykoproteiner kan fungere som slike reseptorer på celleoverflaten. Slike følsomme steder for individuelle stoffer kan spres over overflaten av cellen eller samles i små soner.
Ulike celler av dyreorganismer kan ha forskjellige sett med reseptorer eller ulik følsomhet for samme reseptor.
Rollen til mange cellereseptorer er ikke bare i bindingen av spesifikke stoffer eller evnen til å reagere på fysiske faktorer, men også i overføringen av intercellulære signaler fra overflaten til cellen. For tiden er systemet for signaloverføring til celler ved hjelp av visse hormoner, som inkluderer peptidkjeder, blitt godt studert. Disse hormonene har vist seg å binde seg til spesifikke reseptorer på overflaten av cellens plasmamembran. Reseptorer, etter binding til hormonet, aktiverer et annet protein, som allerede er i den cytoplasmatiske delen av plasmamembranen, adenylatcyklase. Dette enzymet syntetiserer det sykliske AMP-molekylet fra ATP. Rollen til syklisk AMP (cAMP) er at det er en sekundær budbringer - en aktivator av enzymer - kinaser som forårsaker modifikasjoner av andre enzymproteiner. Så når bukspyttkjertelhormonet glukagon, produsert av A-celler på holmene i Langerhans, virker på levercellen, binder hormonet seg til en spesifikk reseptor, som stimulerer aktiveringen av adenylatcyklase. Syntetisert cAMP aktiverer proteinkinase A, som igjen aktiverer en kaskade av enzymer som til slutt bryter ned glykogen (dyrelagringspolysakkarid) til glukose. Virkningen av insulin er motsatt - det stimulerer inngangen av glukose til levercellene og dens avsetning i form av glykogen.
Generelt utfolder hendelseskjeden seg som følger: hormonet interagerer spesifikt med reseptordelen av dette systemet og, uten å trenge inn i cellen, aktiverer adenylatcyklase, som syntetiserer cAMP, som aktiverer eller hemmer et intracellulært enzym eller en gruppe enzymer . Dermed blir kommandoen, signalet fra plasmamembranen overført inne i cellen. Effektiviteten til dette adenylatcyklasesystemet er meget høy. Dermed kan samspillet mellom ett eller flere hormonmolekyler føre, på grunn av syntesen av mange cAMP-molekyler, til en signalforsterkning tusenvis av ganger. I dette tilfellet fungerer adenylatcyklasesystemet som en omformer av eksterne signaler.
Det er en annen måte som andre sekundære budbringere brukes på - dette er den såkalte. fosfatidylinositol-vei. Under påvirkning av et passende signal (noen nervemediatorer og proteiner) aktiveres enzymet fosfolipase C, som spalteripidet, som er en del av plasmamembranen. Hydrolyseproduktene av dette lipidet aktiverer på den ene siden proteinkinase C, som aktiverer kinasekaskaden, som fører til visse cellulære reaksjoner, og på den annen side fører til frigjøring av kalsiumioner, som regulerer en rekke cellulære prosesser.
Et annet eksempel på reseptoraktivitet er reseptorene for acetylkolin, en viktig nevrotransmitter. Acetylkolin, som frigjøres fra nerveenden, binder seg til reseptoren på muskelfiberen, forårsaker en impulsiv strøm av Na + inn i cellen (membrandepolarisering), og åpner umiddelbart rundt 2000 ionekanaler i området av den nevromuskulære enden.
Mangfoldet og spesifisiteten til settene av reseptorer på overflaten av celler fører til dannelsen av et veldig komplekst system av markører som gjør det mulig å skille ens egne celler (av samme individ eller av samme art) fra andres. Lignende celler inngår interaksjoner med hverandre, noe som fører til adhesjon av overflater (konjugering i protozoer og bakterier, dannelse av vevscellekomplekser). I dette tilfellet blir celler som er forskjellige i settet av determinantmarkører eller som ikke oppfatter dem, enten ekskludert fra slik interaksjon, eller blir ødelagt i høyere dyr som et resultat av immunologiske reaksjoner (se nedenfor).
Plasmamembranen er assosiert med lokalisering av spesifikke reseptorer som reagerer på fysiske faktorer. Så i plasmamembranen eller dens derivater i fotosyntetiske bakterier og blågrønne alger, er reseptorproteiner (klorofyller) som interagerer med lyskvanter lokalisert. I plasmamembranen til lysfølsomme dyreceller er det et spesielt system av fotoreseptorproteiner (rhodopsin), ved hjelp av hvilke lyssignalet omdannes til et kjemisk, som igjen fører til generering av en elektrisk impuls.
Intercellulær gjenkjennelse
I flercellede organismer, på grunn av intercellulære interaksjoner, dannes komplekse cellulære ensembler, hvis vedlikehold kan utføres på forskjellige måter. I germinalt, embryonalt vev, spesielt i de tidlige utviklingsstadiene, forblir celler koblet til hverandre på grunn av overflatenes evne til å holde sammen. Denne eiendommen vedheft(tilkobling, adhesjon) av celler kan bestemmes av egenskapene til overflaten deres, som spesifikt samhandler med hverandre. Mekanismen til disse forbindelsene er godt studert, den er gitt av interaksjonen mellom glykoproteiner i plasmamembraner. Med en slik intercellulær interaksjon av celler mellom plasmamembraner forblir det alltid et gap på omtrent 20 nm bredt, fylt med glykokalyx. Behandling av vev med enzymer som krenker integriteten til glykokalyxen (slim som virker hydrolytisk på muciner, mukopolysakkarider) eller skader plasmamembranen (proteaser) fører til isolering av celler fra hverandre, til deres dissosiasjon. Men hvis dissosiasjonsfaktoren fjernes, kan cellene settes sammen igjen og reaggregere. Så det er mulig å dissosiere celler av svamper i forskjellige farger, oransje og gul. Det viste seg at det dannes to typer aggregater i blandingen av disse cellene: de som kun består av gule og kun oransje celler. I dette tilfellet organiserer blandede cellesuspensjoner seg selv, og gjenoppretter den opprinnelige flercellede strukturen. Lignende resultater ble oppnådd med separerte cellesuspensjoner av amfibieembryoer; i dette tilfellet er det en selektiv romlig separasjon av ektodermceller fra endodermen og fra mesenkymet. Videre, hvis vev fra sene stadier av embryonal utvikling brukes til reaggregering, samles ulike celleensembler med vev- og organspesifisitet uavhengig i et reagensrør, epitelaggregater som ligner på nyretubuli, etc.
Det ble funnet at transmembrane glykoproteiner er ansvarlige for aggregeringen av homogene celler. Direkte for forbindelsen, vedheft, celler er ansvarlige for molekylene til den såkalte. CAM-proteiner (celleadhesjonsmolekyler). Noen av dem forbinder celler med hverandre på grunn av intermolekylære interaksjoner, andre danner spesielle intercellulære forbindelser eller kontakter.
Interaksjoner mellom adhesive proteiner kan være homofil når naboceller binder seg til hverandre ved hjelp av homogene molekyler, heterofile når forskjellige typer CAM-er på naboceller er involvert i adhesjon. Intercellulær binding skjer gjennom ytterligere linkermolekyler.
Det finnes flere klasser av CAM-proteiner. Dette er cadheriner, immunglobulinlignende N-CAM (nervecelleadhesjonsmolekyler), selektiner, integriner.
Cadheriner er integrerte fibrillære membranproteiner som danner parallelle homodimerer. Separate domener av disse proteinene er assosiert med Ca 2+ ioner, noe som gir dem en viss stivhet. Det er mer enn 40 arter av cadheriner. Dermed er E-cadherin karakteristisk for celler fra forhåndsimplanterte embryoer og epitelceller fra voksne organismer. P-cadherin er karakteristisk for trofoblast-, placenta- og epidermisceller; N-cadherin er lokalisert på overflaten av nerveceller, linseceller og på hjerte- og skjelettmuskulatur.
Adhesjonsmolekyler for nerveceller(N-CAM) tilhører immunoglobulin-superfamilien, de danner forbindelser mellom nerveceller. Noen av N-CAM-ene er involvert i koblingen av synapser, så vel som i adhesjonen av celler i immunsystemet.
selectins også, integrerte proteiner i plasmamembranen er involvert i adhesjonen av endotelceller, i bindingen av blodplater, leukocytter.
Integriner er heterodimerer, med a- og b-kjeder. Integriner forbinder først og fremst celler med ekstracellulære substrater, men de kan også delta i celleadhesjon til hverandre.
Gjenkjennelse av fremmede proteiner
Som allerede nevnt utvikler fremmede makromolekyler (antigener) som har kommet inn i kroppen en kompleks kompleks reaksjon - en immunreaksjon. Dens essens ligger i det faktum at noen av lymfocyttene produserer spesielle proteiner - antistoffer som spesifikt binder seg til antigener. For eksempel gjenkjenner makrofager antigen-antistoffkomplekser med deres overflatereseptorer og absorberer dem (for eksempel absorpsjon av bakterier under fagocytose).
I kroppen til alle virveldyr er det i tillegg et system for mottak av fremmede celler eller deres egne, men med endrede plasmamembranproteiner, for eksempel under virusinfeksjoner eller mutasjoner, ofte assosiert med tumordegenerasjon av celler.
Proteiner er lokalisert på overflaten av alle virveldyrceller, den såkalte. stort histokompatibilitetskompleks(hoved histokompatibilitetskompleks - MHC). Dette er integrerte proteiner, glykoproteiner, heterodimerer. Det er veldig viktig å huske at hvert individ har et annet sett av disse MHC-proteinene. Dette skyldes det faktum at de er veldig polymorfe, fordi hvert individ har et stort antall alternerende former av samme gen (mer enn 100), i tillegg er det 7-8 loci som koder for MHC-molekyler. Dette fører til det faktum at hver celle i en gitt organisme, som har et sett med MHC-proteiner, vil være forskjellig fra cellene til et individ av samme art. En spesiell form for lymfocytter, T-lymfocytter, gjenkjenner MHC i kroppen, men den minste endring i strukturen til MHC (for eksempel assosiasjon med et virus eller resultatet av en mutasjon i individuelle celler), fører til faktum at T-lymfocytter gjenkjenner slike endrede celler og ødelegger dem, men ikke ved fagocytose. De skiller ut spesifikke perforinproteiner fra sekretoriske vakuoler, som er innebygd i den cytoplasmatiske membranen til den endrede cellen, danner transmembrankanaler i den, noe som gjør plasmamembranen permeabel, noe som fører til at den endrede cellen dør (fig. 143, 144).
Spesielle intercellulære forbindelser
I tillegg til disse relativt enkle adhesive (men spesifikke) bindingene (fig. 145), er det en rekke spesielle intercellulære strukturer, kontakter eller forbindelser som utfører visse funksjoner. Dette er låse-, forankrings- og kommunikasjonsforbindelser (fig. 146).
Låse eller tett forbindelse karakteristisk for enkeltlags epitel. Dette er sonen hvor de ytre lagene av de to plasmamembranene er så nærme som mulig. Trelagsmembranen sees ofte i denne kontakten: de to ytre osmofile lagene til begge membranene ser ut til å smelte sammen til ett felles lag 2–3 nm tykt. Fusjon av membraner skjer ikke over hele området med tett kontakt, men er en serie av punktkonvergens av membraner (fig. 147a, 148).
På plane preparater av plasmamembranbrudd i sonen med tett kontakt, ved bruk av fryse- og flismetoden, ble det funnet at kontaktpunktene til membranene er rader med kuler. Dette er proteinene occludin og claudin, spesielle integrerte proteiner i plasmamembranen, bygget i rader. Slike rader med kuler eller strimler kan krysse hverandre på en slik måte at de så å si danner et gitter eller nettverk på spalteflaten. Denne strukturen er veldig typisk for epitel, spesielt kjertel og tarm. I det siste tilfellet danner tett kontakt en kontinuerlig sone for fusjon av plasmamembraner, som omgir cellen i dens apikale (øvre, ser inn i tarmlumen) del (fig. 148). Dermed er hver celle i laget så å si omgitt av et bånd av denne kontakten. Slike strukturer kan også sees med spesielle flekker i et lysmikroskop. De fikk navnet fra morfologer endeplater. Det viste seg at i dette tilfellet er rollen til den lukkende tette kontakten ikke bare i den mekaniske forbindelsen av celler med hverandre. Dette kontaktområdet er dårlig permeabelt for makromolekyler og ioner, og dermed låser det, blokkerer de intercellulære hulrommene, og isolerer dem (og med dem det indre miljøet i kroppen) fra det ytre miljøet (i dette tilfellet tarmlumen).
Dette kan demonstreres ved bruk av elektrontette kontrastmidler som lantanhydroksidløsning. Hvis lumen i tarmen eller kanalen til en kjertel er fylt med en løsning av lantanhydroksid, så på seksjoner under et elektronmikroskop, har sonene der dette stoffet befinner seg en høy elektrontetthet og vil være mørke. Det viste seg at verken sonen med tett kontakt eller de intercellulære rommene under den mørkere. Hvis tight junctions er skadet (ved lett enzymatisk behandling eller fjerning av Ca ++ ioner), trenger lantan også inn i de intercellulære områdene. Tilsvarende har tight junctions vist seg å være ugjennomtrengelige for hemoglobin og ferritin i tubuli i nyrene.